

 Genetic analysis of cave bear specimens from Niedźwiedzia Cave, Sudetes, Poland
Genetic analysis of cave bear specimens from Niedźwiedzia Cave, Sudetes, Poland
Article number: 15.2.21A
https://doi.org/10.26879/301
Copyright Palaeontological Association, July 2012
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 8 September 2011. Acceptance: 5 June 2012
{flike id=263}
ABSTRACT
The vast majority of fossil remains in Late Pleistocene deposits from Niedźwiedzia Cave in Kletno, Sudetes, Poland, belong to the cave bear. Phylogenetic analyses based on a fragment of the mitochondrial D-loop region extracted from two cave bear samples unambiguously showed their close relationship with the Ursus ingressus haplogroup. This taxonomic affiliation of the cave bear remains from Niedźwiedzia Cave was further confirmed by biometrical analyses of molar teeth and skulls. Our results represent the first record of U. ingressus north of the Carpathian Arch, while radiocarbon dating (> 49,000 yr BP) of the samples indicates that they represent some of the oldest specimens of this cave bear taxon known so far. Multi-method phylogenetic analyses including numerous publicly available cave bear sequences allowed analysing the relationships among these samples in details, including the significance of particular clades, and discussing some aspects of cave bear phylogeography. The sequences of U. ingressus from Poland are most closely related to specimens from the Ural Mountains and next to Slovenia, which may indicate migrations between Central and Eastern European populations. The internal placement of Ural samples among European specimens in phylogenetic trees and the older age of Polish samples than those from Urals suggest that the eastward expansion of U. ingressus may have started from Central Europe.
Mateusz Baca.Center for Precolumbian Studies, University of Warsaw, Krakowskie Przedmieście 26/28, 00-927 Warsaw, Poland. bacamat@gmail.com
Anna Stankovic. Institute of Biochemistry and Biophysics, Polish Academy of Science, Pawińskiego 5a, 02-106 Warsaw, Poland. anams@ibb.waw.pl
Krzysztof Stefaniak. Department of Palaeozoology, Zoological Institute, University of Wrocław, Sienkiewicza 21, 50-335 Wrocław, Poland. stefanik@biol.uni.wroc.pl
Adrian Marciszak. Department of Palaeozoology, Zoological Institute, University of Wrocław, Sienkiewicza 21, 50-335 Wrocław, Poland. caspian8@wp.pl
Michael Hofreiter. Department of Biology (Area 2), The University of York, Wentworth Way, Heslington York, YO10 5DD, United Kingdom. michi@palaeo.eu
Adam Nadachowski. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska 17, 31-016 Cracow, Poland. nadachowski@isez.pan.krakow.pl
Piotr Węgleński. Institute of Biochemistry and Biophysics, Polish Academy of Science, Pawińskiego 5a, 02-106 Warsaw, Poland. wegle@adm.uw.edu.pl
Paweł Mackiewicz. Department of Genomics, Faculty of Biotechnology, University of Wrocław, Przybyszewskiego 63/77, 51-148 Wrocław, Poland. pamac@smorfland.uni.wroc.pl
Keywords: ancient DNA; cave bear; phylogeny; Ursus ingressus; Ursus spelaeus
Final citation: Baca, Mateusz, Stankovic, Anna, Stefaniak, Krzysztof, Marciszak, Adrian, Hofreiter, Michael, Nadachowski, Adam, Węgleński, Piotr, and Mackiewicz, Paweł. 2012. Genetic analysis of cave bear specimens from Niedźwiedzia Cave, Sudetes, Poland. Palaeontologia Electronica Vol. 15, Issue 2;21A,16p;
palaeo-electronica.org/content/2012-issue-2-articles/263-cave-bears-from-poland
INTRODUCTION
Out of the big mammal species that went extinct during the Pleistocene, the cave bear is probably the most thoroughly genetically studied one. Its fossil material is quite abundant in cave sites across Europe and Asia, and in many cases preserved sufficiently well to allow ancient DNA (aDNA) extraction and analysis (e.g., Hänni et al., 1994; Loreille et al., 2001; Hofreiter et al., 2002, 2004a, 2004b, 2007; Orlando et al., 2002; Noonan et al., 2005; Valdiosera et al., 2006; Bon et al., 2008; Krause et al., 2008; Knapp et al., 2009; Stiller et al., 2010). Cave bears diverged from their sister lineage leading to the brown bear (Ursus arctos), approximately 1.2-1.7 million years ago according to palaeontological (Kurtén, 1968, 1976; Rustioni and Mazza, 1992; Rabeder and Withalm, 2006) and molecular data (Loreille et al., 2001; Bon et al., 2008), although some calculations indicate an earlier split about 2.8 million years ago (Krause et al., 2008). It is usually assumed that Late Pleistocene cave bears (Ursus ex gr. spelaeus) evolved gradually from the Middle Pleistocene cave bears from the deningeri-group still before the Last Interglacial (Mazza and Rustioni, 1994; Rabeder et al., 2000, 2010).
The results of both morphological and genetic studies have shown that during the Late Pleistocene, Europe was inhabited by two main cave bear haplogroups described as separate species, U. spelaeus and U. ingressus (Rabeder, 1995; Hofreiter et al., 2004b; Rabeder and Hofreiter, 2004; Rabeder et al., 2004a, 2004b). They separated between 173,000-414,000 yr BP according to molecular dating (Knapp et al., 2009). Additionally, among U. spelaeus several high-alpine forms with reduced body size, named U. s. eremus and U. s. ladinicus, were discerned whereas the 'classic' Western Europe cave bear was called U. s. spelaeus (Rabeder et al., 2000, 2004a, 2004b, 2008; Hofreiter et al., 2004b; Rabeder and Hofreiter, 2004). For a recent review of most described cave bear taxa and a somewhat different conception of their classification based on craniometrical features see Baryshnikov and Puzachenko (2011). According to this study, the morphological differences between U. spelaeus and U. ingressus do not exceed subspecies level and are too small to consider them as different species.
A large number of radiocarbon dates revealed that cave bears became extinct just before the Last Glacial Maximum (LGM), ca. 24,000 yr BP (27,800 cal. yr BP; Pacher and Stuart, 2009). Comparison of effective female population sizes of cave bears and their sister species, the brown bear, showed that the genetic decline of cave bears preceded their extinction by about 25,000 years, while brown bear population size remained constant (Stiller et al., 2010). The reason(s) for the extinction of cave bears are still unclear but it seems that not only one, but probably several causes played a role in its disappearance, possibly including climatic changes, human impact (Stiller et al., 2010), and predator stress caused by cave lion and cave hyena (Diedrich, 2010).
The phylogeographic picture of cave bears in Eurasia does not offer clear indications of the migration routes of cave bears during the Pleistocene glacials and interglacials. It has been suggested that U. ingressus dominated mainly in Eastern and Central Europe while U. spelaeus did so in Western Europe (Hofreiter et al., 2004a; Knapp et al., 2009). In addition to this picture in Europe, remains of both species were also discovered in Asia (Knapp et al., 2009). Whereas U. spelaeus inhabited mainly low and medium elevation areas, U. ingressus has mostly been found in medium and high elevated regions (Baryshnikov and Puzachenko, 2011). Recent isotopic analyses showed also some dietary differentiation between these cave bear haplogroups (Bocherens et al., 2011; Dotsika et al., 2011). Ursus ingressus was likely better adapted to continental environments and, thus, might have outperformed U. spelaeus during cold and arid climate conditions (Baryshnikov and Puzachenko, 2011). In fact, it was shown that U. ingressus immigrated into the Alps around 50,000 years ago where it replaced two former populations of U. s. eremus and U. s. ladinicus (Rabeder and Hofreiter, 2004; Rabeder et al., 2008). Similarly, the stratigraphically younger U. ingressus replaced quite suddenly the older U. s. spelaeus around 28,000 yr BP (32,000 cal. BP) in three geographically close caves in the Swabian Jura of Germany (Münzel et al., 2011). However, despite the numerous samples that have been genetically analysed so far, there are a number of regions from which no genetic data have been obtained so far and which could contribute to a better understanding of the complex picture of recent cave bear evolution.
Although fossil remains of cave bears are quite common in Poland, i.e., north of the Carpathian and Sudetes Mountain ridges, no DNA analyses were so far performed on cave bear remains from this region of Europe. Such analyses would be an important step towards reconstructing a more complete picture of phylogeographic structure and genetic variation of cave bears sensu lato. In this paper we present the results of the analysis of cave bear specimens discovered in Niedźwiedzia Cave (Wiszniowska, 1976; Wiszniowska et al., 1996; Bieroński et al., 2009) located in the eastern Sudetes Mountains. Phylogenetic analyses were performed with a variety of methods to ensure that the obtained tree topology was not dependent on a specific method.
MATERIALS AND METHODS
 Characterization of Niedźwiedzia Cave, Its Deposits and Palaeoenvironment
Characterization of Niedźwiedzia Cave, Its Deposits and Palaeoenvironment
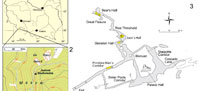
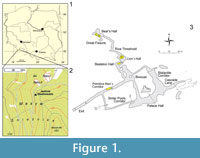
Niedźwiedzia Cave (Polish for Bear Cave) (50° 14' 03'' N; 016° 50' 03'' E) is located in the village of Kletno, Stronie Śląskie commune, in the Śnieźnik Massif of the Sudetes (Figure 1). It was discovered in 1967 during exploitation of crystaline limestone (marble) in a small quarry at the Kleśnica stream valley. Artificial entrances to the cave are situated at 800 and 807 m above sea level. The length of the known passages is more than 3,000 m. A systematic, detailed palaeontological exploration of Niedźwiedzia Cave from 1967 until today yielded a huge collection of a few hundred thousand vertebrate remains and snail shells (Wiszniowska, 1976, 1978, 1989; Wiszniowska et al., 1996; Bieroński et al., 2009). The age of the deposits containing bone debris of Pleistocene animals was estimated as upper Vistula glaciation (> 50,000 – 12,000 yr BP), with Holocene deposits only in the top parts of some profiles (Wiszniowska, 1989; Wiszniowska et al., 1996; Bieroński et al., 2009).
The radiocarbon dating of cave bear remains from different profiles gave ages from 32,100±1,300 to > 49,000 yr BP (Bieroński et al., 2009). All dates correspond to the period MIS 3, from the Grudziądz Interstadial to the main stadial of the Vistula glaciation.
All cave bear remains from diverse localities in Poland date to a period before the LGM as in the rest of Europe. However, the youngest dates from Poland (ca. 26,000 yr BP, cal. 31,000 yr BP) are older than those of cave bears from the Alps, suggesting that in areas north of the Sudetes and the Carpathian ranges cave bears became extinct a bit earlier (Nadachowski et al., 2010).
The fossil faunal assemblage from Niedźwiedzia Cave includes 30 mammal species (Wiszniowska, 1989; Bieroński et al., 2009), many of which live today in the temperate zone and are associated with meadows, tree stands, and forests, such as field vole Microtus arvalis, European water vole Arvicola terrestris, bank vole Myodes glareolus, badger Meles meles, pine marten Martes martes, brown bear Ursus arctos, wild boar Sus scrofa, red deer Cervus elaphus, and roe deer Capreolus capreolus. The remaining species, such as red fox Vulpes vulpes and wolf Canis lupus, are regarded as euryoecious, i.e., adapted to an array of habitats. Extinct mammals are represented by cave bear Ursus ex gr. spelaeus, cave hyena Crocuta crocuta spelaea, cave lion Panthera spelaea, and bison Bison priscus. The remains of the cave bear were the most abundant and constituted from 70 to 90% of the bone material in all profiles. The extinct species were components of the faunal association of the so-called mammoth steppe, characteristic of cold periods of the Pleistocene while the presence of chamois Rupicapra rupicapra remains indicates open areas or pine forest, or alternatively mountain forests on a rocky substratum.
Fossil Material Used in a DNA Extraction
Two fossil specimens of cave bear used in this study, i.e., a small fragment of the crown of one cave bear's canine and a proximal epiphysis of a left ulna bone, were excavated from profiles located in Lion`s Hall and Primitive Man`s Corridor in Niedźwiedzia Cave (Figure 1.3). They were found during the 1967-1972 excavations in cave loam deposit, which for the most part filled the cave's corridors. They do not provide any morphometric diagnostic features to classify them to one of the two Late Pleistocene cave bear taxa U. ingressus or U. spelaeus. It can only be concluded that the relatively small size of the ulna bone (anterio-posterior length ca. 71.0 mm) allows assigning it to a female. The specimens are hosted in the collection of the Palaeozoology Department, Zoological Institute, Wrocław University (ZPALUWr/JN/K). Samples that yielded reliable mtDNA sequences were AMS C14 dated in the Poznan Radiocarbon Laboratory, Poznań, Poland (Table 1). The radiocarbon dates were calibrated using CalPal.
Contamination Precautions
DNA extraction and PCR setup were performed in a laboratory dedicated especially to ancient DNA work with positive air pressure. All staff wore lab coats, face masks and gloves. Working areas and benches were frequently cleaned with bleach and DNA ExitusPlus (Applichem). All the reactions were carried out in laminar flow cabinets with HEPA filters. Only filter tips and sterile disposables were used.
DNA Extraction, Amplification and Sequencing
Prior to DNA extraction each tooth or bone fragment was cleaned with bleach, rinsed with ddH2O, UV irradiated for at least 20 minutes on each side and pulverized in a cryogenic mill (Spex CentriPrep). About 100 mg of powder were incubated overnight at 40°C in 1.6 ml of extraction buffer (0.5 M EDTA, 0.7 mg of proteinase K (Bioline), 0.1 M DTT, 50 mM PTB, 0.5% N-Lauryl sarcosine salt) with constant agitation. The supernatant was subjected to phenol:chloroform:isoamyl alcohol (25:24:1, v:v:v) extraction, followed by two chloroform extractions and isopropanol precipitation (Sambrook et al., 1989). After precipitation, the DNA was resuspended in 60 μl of ddH2O (Fluka). At least two independent extractions were performed from each sample. A 285 bp fragment of mtDNA control region was amplified with three overlapping primer pairs (1F: 5'-CTA TTT AAA CTA TTC CCT GGT ACA TAC-3'; 1R: 5'-ATG GGG GCA CGC CAT TAA TGC-3'; 2F: 5'-CAT CTC ATG TAC TGT ACC ATC ATA GT-3'; 2R: 5'-TAA ACT TTC GAA ATG TAG GTC CTC ATG-3'; 3F: 5'-GCC CCA TGC ATA TAA GCA TG-3'; 3R: 5'-GGA GCG AGA AGA GGT ACA CGT-3'). All primers were taken from Hofreiter et al. (2002) with exception of the primer 3R, which was redesigned. PCR reactions were carried out in 25 μl of reaction mix containing 0.2 μM of each primer, 1X AmpliTaq Gold Master Mix (ABI), 0.1 M BSA and 1.5 μl of extracted DNA. Initial denaturation of 12 min. at 95°C was followed by 40 cycles of 30 sec. at 95 °C, 45 sec. at 41°C, 1 min. at 72°C followed by a final extension of 7 min. at 72°C. PCR products were purified with SureClean Kit (Bioline) and sequenced. To ensure that PCR products were homogenous and the obtained sequences were not erroneous due to post-mortem DNA modifications, each product was cloned with the PCR Cloning Kit (Qiagen) according to the manufacturer's instruction. Several clones were sequenced from each isolation and primer pair and checked for consistency.
For two specimens, CB4 and CB5, a 285 bp fragment of the D-Loop sequence was obtained. The sequences were deposited in GenBank under accession numbers JN112374 and JN112375.
Comparative Biometrical Analysis of Cave Bear Remains
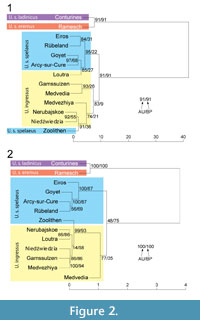
For comparison of the cave bear remains from Niedźwiedzia Cave with other samples, whose taxonomic affiliation was indicated by genetic or morphological studies, we used length and width of all molars published by Baryshnikov (2006), Baryshnikov et al. (2003), Nagel et al. (2005), Rabeder (1995), Rabeder (2004b), Sabol (2005), Tsoukala et al. (2006) and Wiszniowska (1976). Apart from Niedźwiedzia Cave (Poland) this data considered the following localities: Arcy-sur-Cure (France), Conturines Cave (Italy), Cova Eiros (Spain), Gamssulzen Cave (Austria), Goyet (Belgium), Loutra Arideas Bear Cave (Greece), Medvezhiya Cave (Urals, Russia), Medvedia Cave (Slovakia), Nerubajskoe (Ukraine), Ramesch Cave (Austria), Rübeland (Germany), and Zoolithen Cave (Germany). Hierarchical clustering analysis was carried out on mean values assuming UPGMA (Unweighted Pair-Group Method Arithmetic Averages), agglomerative method and squared Euclidean distance measure using the R package (R Development Core Team, 2010). To evaluate the reliability of specific clades in the obtained dendrograms we applied approximately unbiased tests and bootstrap resampling assuming 1000 replications.
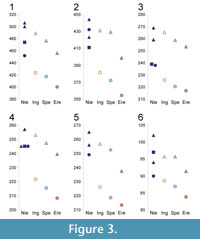
Skull measurements for six cave bears from Niedźwiedzia Cave were taken from Wiszniowska (1976) and compared to corresponding measurements of cave bear taxa by Baryshnikov and Puzachenko (2011).
Phylogenetic Analyses
To determine the phylogenetic relationships of the DNA sequences from Niedźwiedzia Cave, we compared them to cave bear sequences that were gathered by thorough BLAST searches of the GenBank database. Three U. arctos sequences were chosen as outgroup according to Knapp et al. (2009). After exclusion of incomplete sequences we obtained a set of 70 sequences covering the part of the mitochondrial D-loop region analysed in this study. Next, we eliminated redundant sequences from this set leaving only one representative for each haplotype. However, all locations of each sequence were included in the final tree. After removal of an ambiguously alignable long pyrimidine stretch, a final set of 39 aligned sequences with 254 bp length was used in all subsequent analyses.
Phylogenetic trees were inferred using 10 approaches. Three Bayesian analyses were done using PhyloBayes 3.2f (Lartillot and Philippe, 2004) and one using MrBayes 3.1.2 (Ronquist and Huelsenbeck, 2003). Maximum likelihood (ML) trees were constructed with PhyMl 3.0 (Guindon and Gascuel, 2003) and PAUP* 4.0b (Swofford, 1998). Additionally, we applied maximum parsimony (MP) and three distance methods: neighbor joining (NJ), minimum evolution (ME) and weighted least squares (WLS). All trees were reconstructed with PAUP. The best-fit substitution model HKY+I as estimated in jModeltest 0.1.1 (Posada, 2008) according to AIC, AICc and BIC criteria, was used with MrBayes, ML, NJ, ME and WLS.
For the PhyloBayes analyses, we applied three of the available models: Poisson (i.e., F81), GTR and the CAT-Poisson model with the number of components, weights and profiles inferred from the data. Two independent Markov chains were run for 1,000,000 generations in each of these analyses. The last 250,000 trees from each chain were collected to compute posterior consensus trees after reaching convergence (i.e., the largest discrepancy across all bipartitions was lower than 0.015, much below the proposed threshold of 0.1). For the MrBayes analyses, two independent runs starting from random trees using four Markov chains each were carried out. Trees were sampled every 100 generations for 30,000,000 generations. In the final analysis we selected trees from the last 6,650,000 generations that reached the stationary phase and convergence (i.e., the standard deviation of split frequencies stabilized and was lower than 0.0014, much below the proposed threshold of 0.01).
We used two heuristic search algorithms in PhyMl, i.e., NNI and SPR. Node support was assessed by non-parametric bootstrap analysis with 1000 replicates and by the approximate likelihood ratio test (aLRT) based on the ![]() 2 and the Shimodaira-Hasegawa-like procedures (Guindon and Gascuel, 2003). The minimum of the support values from the latter two procedures was shown at nodes in the presented tree. In the case of ML, ME, WLS and MP methods used with PAUP, final trees were searched from 10 starting trees obtained by stepwise addition with random-addition sequence. Non-parametric bootstrap analyses were performed with 1000 replicates for each reconstruction done with PAUP. The tree-bisection-reconnection (TBR) branch-swapping algorithm was applied in the final tree search and bootstrap procedures for PAUP ML, ME, WLS and MP methods.
2 and the Shimodaira-Hasegawa-like procedures (Guindon and Gascuel, 2003). The minimum of the support values from the latter two procedures was shown at nodes in the presented tree. In the case of ML, ME, WLS and MP methods used with PAUP, final trees were searched from 10 starting trees obtained by stepwise addition with random-addition sequence. Non-parametric bootstrap analyses were performed with 1000 replicates for each reconstruction done with PAUP. The tree-bisection-reconnection (TBR) branch-swapping algorithm was applied in the final tree search and bootstrap procedures for PAUP ML, ME, WLS and MP methods.
The reliability of clades in the resulting tree was also evaluated by the weighted least-squares likelihood ratio test (WLS-LRT) and F-test as implemented in the WeightLESS 3.05 program (Sanjuan and Wrobel, 2005). The distance matrix for the original sequences and 1000 bootstrap distance matrices used in this program were calculated with PAUP assuming the HKY+I model.
Tree topologies obtained by all 10 approaches were compared according to the approximately unbiased test (AU) with the Consel v0.1k program (Shimodaira and Hasegawa, 2001) with 10,000,000 replicates. Site-wise log-likelihoods for the analysed trees were calculated in PAUP under the HKY+I model.
In addition to the phylogenetic analyses, a median-joining network (Bandelt et al., 1999) based on the 254 bp alignment of all 70 cave bear sequences was constructed using the Network 4.6 software (fluxus-engineering.com). The Maximum Parsimony algorithm was used to resolve reticulations in the final network (Polzin and Daneschmand, 2003).
RESULTS AND DISCUSSION
 Taxonomic Affiliation of Cave Bear Remains from Niedźwiedzia Cave Based on Biometrical Studies
Taxonomic Affiliation of Cave Bear Remains from Niedźwiedzia Cave Based on Biometrical Studies
Although it is possible to discriminate U. ingressus from high-alpine forms of U. spelaeus by dental morphometry and long bones measurements (Rabeder, 2004; Rabeder and Hofreiter, 2004: Rabeder et al., 2004b, 2008), the morphological differences between U. ingressus and U. s. spelaeus are much less pronounced (Münzel et al., 2009, 2010; Baryshnikov and Puzachenko, 2011). Nevertheless, we compared length and width of molars (Figure 2) as well as main skull measurements (Figure 3) of cave bear specimens from Niedźwiedzia Cave with other remains whose taxonomic affiliation was confirmed by genetic or morphological studies.
 Molars from Niedźwiedzia Cave are much larger than those from Conturines and Ramesch caves, and in a clustering analysis, they group with specimens assigned to U. ingressus with statistically significant support (Figure 2). Interestingly, this analysis also showed a clear separation of most analysed representatives of the two large cave bear forms U. ingressus and U. s. spelaeus into two clades. Only teeth from Zoolithen Cave, a locality typical of U. s. spelaeus, were usually clustered with U. ingressus. However, these results may be biased by an unbalanced sex ratio of the teeth sets used and a mixture of teeth coming from different cave bear taxa. For example, genetic analyses of Zoolithen Cave samples revealed the presence of mtDNA typical of not only U. s. spelaeus but also U. ingressus in this cave (Stiller et al., 2009).
Molars from Niedźwiedzia Cave are much larger than those from Conturines and Ramesch caves, and in a clustering analysis, they group with specimens assigned to U. ingressus with statistically significant support (Figure 2). Interestingly, this analysis also showed a clear separation of most analysed representatives of the two large cave bear forms U. ingressus and U. s. spelaeus into two clades. Only teeth from Zoolithen Cave, a locality typical of U. s. spelaeus, were usually clustered with U. ingressus. However, these results may be biased by an unbalanced sex ratio of the teeth sets used and a mixture of teeth coming from different cave bear taxa. For example, genetic analyses of Zoolithen Cave samples revealed the presence of mtDNA typical of not only U. s. spelaeus but also U. ingressus in this cave (Stiller et al., 2009).
The comparison of available skull measurements including sexual dimorphism also revealed closer similarity of the remains from Niedźwiedzia Cave to U. ingressus (Figure 3). The medians of skull measurements calculated by Baryshnikov and Puzachenko (2011) showed an increasing trend from U. s. eremus, through U. s. spelaeus to U. ingressus. The skull measurements of the Niedzwiedzia Cave specimens fit well into this trend and generally resemble the ones of U. ingressus skulls.
Dating of Cave Bear Remains from Niedźwiedzia Cave
The sample CB4 was AMS C14 dated to 41,500±1,100 yr BP, while sample CB5 was dated to > 49,000 yr BP (Table 1). The most recent AMS radiocarbon dating of other cave bear remains from Niedźwiedzia Cave also indicated similar ages from 41,300±1,300 to > 49,000 yr BP. Thus, both samples analysed in this work belong to the oldest U. ingressus specimens described so far (Table 2).
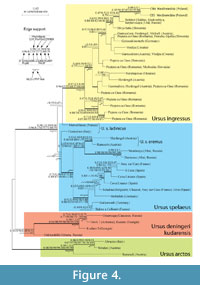 Phylogenetic Analyses of mtDNA from Cave Bears
Phylogenetic Analyses of mtDNA from Cave Bears
For construction of phylogenetic trees a 254 bp long D-loop alignment of 39 cave bear sequences was used. Trees obtained with 10 approaches gave almost identical or very similar topologies (Figure 4). The AU test did not show statistically significant differences between them.
Similarly to the results obtained by Knapp et al., (2009), several main well-supported clades of cave bears can be recognized in our phylogenies (Figure 4). One clade includes sequences assigned to U. deningeri kudarensis from the Caucasus and Siberia (Yana River) (Knapp et al., 2009). The Siberian sample occupies a position basal to a strongly supported subclade of cave bear sequences originating from the Caucasus. As a monophyletic clade, the U. deningeri kudarensis group is, however, strongly and moderately supported only by the two PhyloBayes approaches. Thus, the Caucasus clade alone is strongly supported, but the position of the Siberian sequence is rather unstable.
Statistical significance in all applied tests, strong support with Bayesian methods (> 0.95) and moderate support (72-81%) in all other analyses were obtained for the second large clade that groups sequences from 'advanced' cave bears, U. spelaeus and U. ingressus. Early branches in this clade are created by two sequences representing alpine forms with reduced body size (named U. s. ladinicus) that were discovered in Merveilleuse and Conturines caves. However, their position in the trees is very poorly supported. Although the sequence from Conturines Cave is grouped in all trees with U. spelaeus sequences, most approaches show an affiliation of the sequence from Merveilleuse Cave with U. ingressus and only in distance trees (LS, ME and NJ) it is placed basal to the rest U. spelaeus sequences. The latter group of sequences is well-supported by Bayesian methods (0.75-0.99) and F-test at the 0.001 level. This group can be further divided into two lineages. The first contains samples from Southwestern and Western Europe whereas the second consists of sequences from the Altai, which cluster with specimens from Austrian medium-sized cave bears classified as U. s. eremus.
The basal positions of polyphyletic lineages of small bears to the large ones suggest that body mass increased independently in several lineages during the evolution of cave bears (Hofreiter et al., 2002). Further supporting this view, recent analyses of mtDNA from another small cave bear form, U. rossicus found in Kizel Cave in the Urals, places this species close to U. ingressus (Münzel et al., 2010: Pacher et al., 2009). These results together with morphological analyses by Baryshnikov and Puzachenko (2011) that demonstrated substantial similarity of U. ingressus to cave bears described as U. kanivetz from the Volga River region, suggest that U. ingressus had likely an East-European origin.
The subclade of the second 'advanced' cave bear species, U. ingressus is strongly supported by aLRT (0.96) and Bayesian methods (≥ 0.95) but only moderately by others (57-64%). However, the WLS-LRT and F-tests unambiguously confirm the statistical significance of this subclade at the 0.001 level. This subclade is further divided into two branches. The one groups four samples from Peştera cu Oase Cave, which are supported by ≥ 0.95 posterior probabilities, 59-76% bootstrap values and the two applied tests. The second branch supported with generally low bootstrap and support values contains samples from both Peştera cu Oase Cave and all other U. ingressus sequences. This tree topology suggests that the Southeastern part of Europe was the place of origin for U. ingressus from which it expanded to other territories in Europe. However, with one exception (see below) the sequences of U. ingressus originating from other countries do not cluster according to their geographic location. Therefore, we cannot infer particular migration routes from the current data.
Both specimens from Niedźwiedzia Cave evidently branch within the U. ingressus subclade and significantly cluster together (posterior probabilities ≥ 0.99; bootstrap values 88-92%, aLRT = 0.92, p-values < 0.05 and < 0.001). These specimens are most closely related to samples that originate from the Urals (Bolshoi Glukhoi, Medvezhiya and Serpievskaya caves). Although the support for this grouping is weak, nine of ten reconstructed trees recovered this relationship. Only the ME tree showed a close relationship between the samples from Niedźwiedzia Cave and the sequence from Divje babe (Slovenia) but with non-significant support. In addition, this tree was marginally rejected by the AU test (with p-value = 0.05). This result indicates that a grouping of the specimens from Niedźwiedzia with those from the Ural caves is more likely than one with the Divje babe (Slovenia) sequence. In agreement with this conclusion, clustering analyses of molar measurements also show a close relationship of Niedźwiedzia and Medvezhiya caves (Figure 2). All sequences from Poland, Urals and Slovenia fall into a distinct clade obtained in all 10 approaches. Bootstrap values of this grouping are slightly lower than 50% but Bayesian support reaches 0.95 and F-test branch significance is at the 0.01 level.
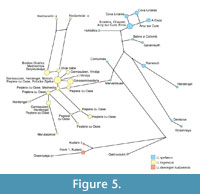 The median joining network shows a similar position of the specimens from Niedźwiedzia Cave (Figure 5). There are also clearly separated groups of U. spelaeus, U. ingressus and U. deningeri kudarensis with U. s. ladinicus in the middle of the network. The haplotypes from Niedźwiedzia Cave are situated in the U. ingressus group and are connected to haplotypes from the Urals and Divje babe.
The median joining network shows a similar position of the specimens from Niedźwiedzia Cave (Figure 5). There are also clearly separated groups of U. spelaeus, U. ingressus and U. deningeri kudarensis with U. s. ladinicus in the middle of the network. The haplotypes from Niedźwiedzia Cave are situated in the U. ingressus group and are connected to haplotypes from the Urals and Divje babe.
The grouping of the Ural specimens with those from Poland may reflect more frequent or recent migrations between Central Europe and the Ural Mountains populations across the East European Plain. The placement of sequences from Ural bears within European samples (see also Stiller et al., 2009) suggests that long-distance migrations were probably directed from Central Europe to the East. Although based on a limited number of samples, this conclusion agrees well with the comparatively old dates obtained for the Polish specimens, which seem to be older than those from the Ural caves. The dates for samples from Niedźwiedzia Cave are 41,500±1,100 and > 49,000 yr BP, whereas a sample from Serpievskaya Cave analysed for DNA has been dated to 44,050 yr BP (Stiller et al., 2010) and the oldest date reported by Kosintev (2007) for mammal bones from Bolshoi Glukhoi is 38,200±900 yr BP. More generally, the oldest published remains of cave bears (including the U. ingressus type) found in the Ural Mountains, from Tayn (Secrets) Cave, do so far not exceed 47,600±900 yr BP (Pacher and Stuart, 2009; Pacher et al., 2009).
This dating indicates that the migration of European cave bears to the Urals may have taken place during Early or Middle Weichselian/Valdai warming periods, e.g., during the Odderade (MIS5a) or Oerel Interstadials (early MIS3). The Odderade Interstadial correlates with the Gniew, Krastudy or Rudunki Interstadials in Polish stratigraphy (Krzywicki, 2002) and with the Krugitsa Interstadial in East European stratigraphy (Velichko and Spasskaya, 2002), whereas the Oerel Interstadial corresponds to the beginning of the Grudziądz Interstadial in Polish stratigraphy (Krzywicki, 2002) and the Krasnogorsk warming of the Bryansk or Grazhdanski megainterval in East European stratigraphy (Velichko et al., 2010).
However, it cannot be excluded that the migration was driven by the asymmetric development of Eurasian ice sheets (Svendsen et al., 2004) and the resulting differences in climatic and environmental conditions between Central and East Europe. For example, during the Middle Weichselian glacial maximum (MIS4) the Barents-Kara Ice Sheet terminated on the northern margin of the Eurasian mainland, whereas the Scandinavian Ice Sheet covered the whole of Finland, reached Denmark and the north-eastern part of Poland. In Polish territory it is known as Świecie or Vistulian 3 Stadial (Krzywicki, 2002), which corresponds to Schalkholz in North Europe and the Shestikhino Stadial in Eastern European stratigraphy (Velichko and Spasskaya, 2002). A more advanced expansion of Ice Sheets in Central Europe could have forced emigrations of cave bears toward the East European Plain, which was free of ice sheets. This region was covered by northern taiga and mixed forests during the Krugitsa and Krasnogorsk Interstadials whereas during the Shestikhino Stadial, the area was covered by periglacial forest-steppe (Velichko et al., 2004). However, more dated DNA samples especially from the East European Plain region are required to support this hypothesis. Such analyses may well reveal more complexity of both the genetic diversity of cave bears and their migration patterns, e.g., in two directions.
Genetic analyses of small cave bears from Ramesch Cave and larger ones from Gamssulzen Cave showed a lack of gene flow between these populations, which inhabited the same alpine region during at least 15,000 years (Hofreiter et al., 2004b). On the other hand, it was found that differences between mtDNA correlate neither with geographical distance nor with the age of samples (Hofreiter et al., 2002) and particular cave bear haplogroups are spread over wide geographic areas (Orlando et al., 2002). Moreover, clustering in phylogenetic trees was observed between geographically distant samples from U. deningeri kudarensis (Caucasus and Yana River) and U. spelaeus (Altai and Austria) (Knapp et al., 2009). Baryshnikov et al., (2003) did not find important morphometrical differences in cave bear cheek teeth from different localities, which they explained by exchange of genetic material between adjacent cave bear populations, whereas they justified the discrepancy between samples from northern and southern sites by different dietary habits. Altogether, the data indicate that long distance migrations occurred in cave bear evolutionary history. Such a view is also supported by the close relationships between the mtDNA sequences from Poland and the Ural, respectively. A specialization and dependence of cave bears on vegetarian diet (see Pacher and Stuart, 2009; Mackiewicz et al., 2010; Bocherens et al., 2011; Dotsika et al., 2011; and references therein) could have been behind this pattern, by forcing cave bear populations to migrate into regions rich enough in food.
CONCLUSIONS
Genetic analyses of mtDNA extracted from cave bear remains found in Niedźwiedzia Cave in Poland clearly indicate its affiliation with the U. ingressus haplogroup. This conclusion is further supported by biometrical analyses of molars and skull. For the first time remains of U. ingressus are identified in the territory of Poland, north of the Carpathians and Sudetes. The investigated samples from Niedźwiedzia Cave belong to one of the oldest recognized remains of this species in Europe. A close relationship of samples from Niedźwiedzia Cave with those from Ural caves, and their respective radiocarbon dates may indicate that migrations or gene flow occurred between Polish cave bear populations and those from the Ural Mountains. These results suggest the existence of a migratory corridor connecting Western and Eastern Europe across the area where Poland is situated today.
ACKNOWLEDGMENTS
This work was supported by Operational Programme Innovative Economy POIG.02.02.00-14-024/08-00.
REFERENCES
Bandelt, H.-J., Forster, P., and Röhl, A. 1999. Median-joining networks for inferring intraspecific phylogenies. Molecular Biology and Evolution, 16:37 – 48.
Baryshnikov, G. 2006. Morphometrical variability of cheek teeth of cave bears, p. 81 – 102. In Scientific Annals, Vol. 98. School of Geology, Aristotle University of Thessaloniki.
Baryshnikov, G. and Puzachenko, A.Y. 2011. Craniometrical variability in the cave bears (Carnivora, Ursidae): Multivariate comparative analysis. Quaternary International, 245:350 – 368.
Baryshnikov, G., Germonpré, M., and Sablin, M. 2003. Sexual dimorphism and morphometric variability of cheek teeth of the cave bears (Ursus spelaeus). Belgian Journal of Zoology, 133:111 –119.
Bieroński , J., Stefaniak, K., Hercman, H., Socha, P., and Nadacowski, A. 2009. Palaeogeographic and palaeoecological analysis of sediments of the Niedźwiedzia Cave in Kletno, p. 401 – 422. In Stefaniak, K., Tyc, A., and Socha, P. (eds.), Karst of the Częstochowa Upland and of the Eastern Sudetes: palaeoenvironments and protection. Studies of the Faculty of Earth Sciences, University of Silesia, No. 56, Sosnowiec-Wrocław.
Bocherens, H., Stiller, M., Hobson, K.A., Pacher, M., Rabeder, G., Burns, J.A., Tütken, T., and Hofreiter, M. 2011. Niche partitioning between two sympatric genetically distinct cave bears (Ursus spelaeus and Ursus ingressus) and brown bear (Ursus arctos) from Austria: isotopic evidence from fossil bones. Quaternary International, 245:238 – 248.
Bon, C., Caudy, N., de Dieuleveult, M., Fosse, P., Philippe, M., Maksud, F., Beraud-Colomb, E., Bouzaid, E., Kefi, R., Laugier, C., Rousseau, B., Casane, D., van der Plicht, J., and Elalouf, J.M. 2008. Deciphering the complete mitochondrial genome and phylogeny of the extinct cave bear in the Paleolithic painted cave of Chauvet. Proceedings of the National Academy of Sciences USA, 105:17447 – 17452.
Diedrich, C.G. 2010. Spotted hyena and steppe lion predation behaviours on cave bears of Europe – Late Quaternary cave bear extinction as result of predator stress. Geophysical Research Abstracts, 12.
Dotsika, E., Zisi, N., Tsoukala, E., Poutoukis, D., Lykoudis, S., and Giannakopoulos, A. 2011. Palaeo-climatic information from isotopic signatures of Late Pleistocene Ursus ingressus bone and teeth apatite (Loutra Arideas Cave, Macedonia, Greece). Quaternary International, 245:291 – 301.
Guindon, S. and Gascuel, O. 2003. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Systematic Biology, 52:696 – 704.
Hänni, C., Laudet, V., Stehelin, D., and Taberlet, P. 1994. Tracking the origins of the cave bear (Ursus spelaeus) by mitochondrial-DNA sequencing. Proceedings of the National Academy of Sciences USA, 91:12336 –12340.
Hofreiter, M., Münzel, S., Conard, N.J., Pollack, J., Slatkin, M., Weiss, G., and Pääbo, S. 2007. Sudden replacement of cave bear mitochondrial DNA in the late Pleistocene. Current Biology, 17:R122 – R123.
Hofreiter, M., Rabeder, G., Jaenicke-Després, V., Withalm, G., Nagel, D., Paunovic, M., Jambrěsić, G., and Pääbo, S. 2004b. Evidence for reproductive isolation between cave bear populations. Current Biology, 14:40 – 43.
Hofreiter, M., Serre, D., Rohland, N., Rebeder, G., Nagel, D., Conrad, N., Münzel, S., and Pääbo, S. 2004a. Lack of phylogeography in European mammals before the last glaciation. Proceedings of the National Academy of Sciences USA, 101:12963 – 12968.
Hofreiter, M., Apelli, C., Krings, M., Waits, L., Conrad, N., Münzel, S., Rabeder, G., Nagel, D., Paunovic, M., Jambrěsić, G., Meyer, S., Weiss, G., and Pääbo, S. 2002. Ancient DNA analyses reveal high mitochondrial DNA sequence diversity and parallel morphological evolution of Late Pleistocene cave bears. Molecular Biology and Evolution, 19:1244 – 1250.
Knapp, M., Rohland, N., Weinstock, J., Baryshnikov, G., Sher, A., Nagel, D., Rabeder, G., Pinhasi, R., Schmidt, H.A., and Hofreiter, M. 2009. First DNA sequences from Asian cave bear fossils reveal deep divergences and complex phylogeographic patterns. Molecular Ecology, 18:1225 – 1238.
Kosintsev, P. 2007. Late Pleistocene large mammal faunas from the Urals. Quaternary International, 160:112 – 120.
Krause, J., Unger, T., Noçon, A., Malaspinas, A.S., Kolokotronis, S.O., Stiller, M., Soibelzon, L., Spriggs, H., Dear, P.H., Briggs, A.W., Bray, S.C., O'Brien, S.J., Rabeder, G., Matheus, P., Cooper, A., Slatkin, M., Pääbo, S., and Hofreiter, M. 2008. Mitochondrial genomes reveal an explosive radiation of extinct and extant bears near the Miocene-Pliocene boundary. BMC Evolutionary Biology, 8:220.
Krzywicki, T. 2002. The maximum ice sheet limit of the Vistulian Glaciation in northeastern Poland and neighbouring areas. Geological Quarterly, 46:165 – 188.
Kurtén, B. 1968. Pleistocene Mammals of Europe. Aldine Publishing, Chicago.
Kurtén, B. 1976. The Cave Bear Story. Life and Death of a Vanished Animal. Columbia University Press, New York.
Lartillot, N. and Philippe, H. 2004. A Bayesian mixture model for across site heterogeneities in the aminoacid replacement process. Molecular Biology and Evolution, 21:1095 – 1109.
Loreille, O., Orlando, L., Patou-Mathis, M., Philippe, M., Taberlet, P., and Hänni, C. 2001. Ancient DNA analysis reveals divergence of the cave bear, Ursus spelaeus, and brown bear, Ursus arctos, lineages. Current Biology, 11:200 – 203.
Mackiewicz, P., Wiszniowska, T., Olejniczak, A.J., Stefaniak, K., Socha, P., and Nadachowski, A. 2010. Analysis of dental enamel thickness in bears with special attention to Ursus spelaeus and U. wenzensis (=minimus) in comparison to selected representatives of mammals, p. 60 – 77. In Nowakowski, D. (ed.), Morphology and Systematics of Fossil Vertebrates. DN Publisher, Wrocław.
Mazza, P. and Rustioni, M. 1994. On the phylogeny of Eurasian bears. Paleontographica Abteilung A, 230:1 – 38.
Münzel, S.C., Pacher, M., and Athen, K. 2009. Correlating genetic results with biometric analysis on metapodial bones. Slovenský Kras Acta Carsologica Slovaca, 47, Supp. l. 1:47 – 56.
Münzel, S.C., Pacher, M., and Athen, K. 2010. Genetic results versus morphological and biometrical analysis - the Case of Ursus spealaeus (sensu lato). 11th ICAZ International Conference, Paris, 23-28 August 2010, Abstracts of the oral and poster presentations, p. 193.
Münzel, S.C., Stiller, M., Hofreiter, M., Mittnik, A., Conard, N.J., and Bocherens, H. 2011. Pleistocene bears in the Swabian Jura (Germany): Genetic replacement, ecological displacement, extinctions and survival. Quaternary International, 245:225 – 237.
Nadachowski, A., Lipecki, G., Stefaniak, K., and Wojtal, P. 2010. Radiocarbon dates on cave bear (Ursus spelaeus) and brown bear (Ursus arctos) from Late Pleistocene of Poland, p. 24. In Bocherens, H. and Pacher, M. (eds.), SSP-4.4 – Late Quaternary mammal ecology: insight from new approaches (direct dating, stable isotopes, DNA). European Geosciences Union. General Assembly 2010, Vienna, Austria, 02-07 May 2010.
Nagel, D., Pronin, K., Rabeder, G., Hofreiter, M., Huijer, W., Kavcik, N., Urbanek, C., Withalm, G., and Orlov, N. 2005. Nerubajskoe, a New Cave Bear Site in the Old Nordmann Territory. Mitteilungen der Kommission für Quartärforschung Österreichische Akademie der Wissenschaften, 14:123 – 134.
Noonan, J.P., Hofreiter, M., Smith, D., Priest, J.R., Rohland, N., Rabeder, G., Krause, J., Detter, J.C., Pääbo, S., and Rubin, E.M. 2005. Genomic sequencing of Pleistocene cave bears. Science, 309:597 – 599.
Orlando, L., Bonjean, D., Bocherens, H., Thenot, A., Argant, A., Otte M., and Hänni, C. 2002. Ancient DNA and the population genetics of cave bears (Ursus spelaeus) through space and time. Molecular Biology and Evolution, 19:1920 – 1933.
Pacher, M. and Stuart, A.J. 2009. Extinction chronology and palaeobiology of the cave bear (Ursus spelaeus). Boreas, 38:189 – 206.
Pacher, M., Stuart, A.J., Baryshnikov, G., Stiller, M., Kosintsev, P., and Vorobiev, A. 2009. Cave bears of the Ural Mountains – a survey based on direct radiocarbon dates, aDNA and morphometrical analysis. 15th International Cave Bear Symposium – Spišská Nová Ves, Slovakia, 17th – 20th of September 2009, Abstract Book, p. 14 – 17.
Polzin, T. and Daneschmand, S.V. 2003. On Steiner trees and minimum spanning trees in hypergraphs. Operations Research Letters, 31:12 – 20.
Posada, D. 2008. jModelTest: phylogenetic model averaging. Molecular Biology and Evolution, 25:1253 – 1256.
R Development Core Team. 2010. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, URL http://www.R-project.org
Rabeder, G. 1995. Evolutionsniveau und Chronologie der Höhlenbären aus der Gamsulzen-Höhle im Toten Gebirge (Oberösterreich). Mitteilungen der Kommission für Quartärforschung Österreichische Akademie der Wissenschaften, 9:69 – 81.
Rabeder, G. 2004. Die Höhlenbären der Sulzfluh-Höhlen. Vorarlberger Naturschau, 15:103 – 114.
Rabeder, G. and Hofreiter, M. 2004. Der neue Stammbaum der Höhlenbären. Die Höhle, 55:1 – 19.
Rabeder, G. and Withalm, G. 2006. Brown bear remains (Ursidae, Mammalia) from Early Pleistocene cave fillings of Deutsch-Altenburg (Lower Austria). 12th International Cave Bear Symposium Aridea/Loutrá, Macedonia, Greece, Greece: School of Geology, Aristotle University of Thessaloniki, Abstract Book, p. 47 – 48.
Rabeder, G., Hofreiter, M., and Withalm, G. 2004a. The systematic position of the cave bear from Potoćka zijalka (Slovenia). Mitteilungen der Kommission für Quartärforschung Österreichische Akademie der Wissenschaften, 13:197 – 200.
Rabeder, G., Nagel, D., and Pacher, M. 2000. Der Höhlenbär. Jan Thorbecke Verlag, Stuttgart.
Rabeder, G., Pacher, M., and Withalm, G. 2010. Early Pleistocene bear remains from Deutsch-Altenburg (Lower Austria). Mitteilungen der Kommission für Quartärforschung Österreichische Akademie der Wissenschaften, Band 17, Vienna.
Rabeder, G., Debeljak, I., Hofreiter, M., and Withalm, G. 2008. Morphological responses of cave bears (Ursus spelaeus group) to high-alpine habitats. Die Höhle, 59:59 – 72.
Rabeder, G., Hofreiter, M., Nagel, D., and Withalm, G. 2004b. New taxa of a cave bears (Ursidae, Carnivora). Cahiers Scientifiques , Hors série, 2:49 – 68.
Ronquist, F. and Huelsenbeck, J.P. 2003. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics, 19:1572 – 1574.
Rustioni, M. and Mazza, P. 1992. The genus Ursus in Eurasia: Dispersal events and stratigraphical significance. Rivista Italiana di Paleontologia e Stratigrafia, 98:487 – 494.
Sabol, M. 2005. Cave Bears (Ursidae, Mammalia) from the Trojuholník Cave (Slovakia). Mitteilungen der Kommission für Quartärforschung Österreichische Akademie der Wissenschaften, 14:161 – 175.
Sambrook, J., Frisch, E.F., and Maniatis, T. 1989. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Lab. Press, Plainview, New York.
Sanjuan, R. and Wrobel, B. 2005. Weighted least-squares likelihood ratio test for branch testing in phylogenies reconstructed from distance measures. Systematic Biology, 54:218 – 229.
Shimodaira, H. and Hasegawa, M. 2001. CONSEL: for assessing the confidence of phylogenetic tree selection. Bioinformatics, 17:1246 – 1247.
Stiller, M., Knapp, M., Stenzel, U., Hofreiter, M., and Meyer, M. 2009. Direct multiplex sequencing (DMPS) - a novel method for targeted high-throughput sequencing of ancient and highly degraded DNA. Genome Research, 19:1843 – 1848.
Stiller, M., Baryshnikov, G., Bocherens, H., Grandal d'Anglade, A., Hilpert, B., Münzel, S.C., Pinhasi, R., Rabeder, G., Rosendahl, W., Trinkaus, E., Hofreiter, M., and Knapp, M. 2010. Withering away - 25,000 years of genetic decline preceded cave bear extinction. Molecular Biology and Evolution, 27:975 – 978.
Svendsen, J.I., Alexanderson, H., Astakhov, V.I., Demidov, I., Dowdeswell, J.A., Funder, S., Gataullin, V., Henriksen, M., Hjort, C., Houmark-Nielsen, M., Hubberten, H.W., Ingólfsson, Ó., Jakobsson, M., Kjær, K.H., Larsen, E., Lokrantz, H., Lunkka, J.P., Lysaa, A., Mangerud, J., Matiouchkov, A., Murray, A., Moeller, P., Niessen, F., Nikolskaya, O., Polyak, L., Saarnisto, M., Siegert, C., Siegert, M.J., Spielhagen, R.F., and Stein, R. 2004. Late Quaternary ice sheet history of northern Eurasia. Quaternary Science Reviews, 23:1229 – 1271.
Swofford, D.L. 1998. PAUP*. Phylogenetic analysis using parsimony (and other methods). Version 4. Sinauer Associates, Sunderland, Massachusetts.
Tsoukala, E., Chatzopoulou, K., Rabeder, G., Pappa, S., Nagel, D., and Withalm, G. 2006. Paleontological and stratigraphical research in Loutra Arideas Bear Cave (Almopia Speleopark, Pella, Macedonia, Greece). Scientific Annals. School of Geology. Aristotle University of Thessaloniki, 98:41 – 67.
Valdiosera, C., García, N., Dalén, L., Smith, C., Kahlke, R.D., Lidén, K., Angerbjörn, A., Arsuaga, J.L., and Götherström, A. 2006. Typing single polymorphic nucleotides in mitochondrial DNA as a way to access Middle Pleistocene DNA. Biology Letters, 2:601 – 603.
Velichko, A. and Spasskaya, I. 2002. Climatic change and the development of landscapes, p. 36 – 69. In Shahgedanova, M. (ed.), The Physical Geography of Northern Eurasia. Oxford University Press, New York.
Velichko, A.A., Faustova, M.A., Gribchenko, Yu N., Pisareva, V., and Sudakova, N.G. 2004. Glaciations of the East European Plain – distribution and chronology, p. 337 – 354. In Ehlers, J. and Gibbard, P.L. (eds.), Quaternary Glaciations — Extent and Chronology. Part I: Europe. Developments in Quaternary Science 2, Elsevier Science.
Velichko, A., Pisareva, V., Morozova, T., Borisova, O., Faustova, M., Gribchenko, Y., Timireva, S., Semenov, V., and Nechaev, V. 2010. Correlation of the glacial and periglacial Pleistocene events in Eastern Europe: lines of attack, p. 188 – 191. In Titov, V.V. and Tesakov, A.S. (eds.), Quaternary stratigraphy and paleontology of the Southern Russia: connections between Europe, Africa and Asia: Abstracts of the International INQUA-SEQS Conference, Rostov-on-Don, June 21–26, 2010.
Wiszniowska, T. 1976. Niedźwiedź jaskiniowy z Kletna i innych jaskiń Polski. Acta Universitatis Wratislaviensis, 311:1 – 75.
Wiszniowska. T. 1978. Panthera spelaea (Goldfuss) z Jaskini Niedźwiedziej w Kletnie. Acta Universitatis Wratislaviensis, 329:113 – 141.
Wiszniowska, T. 1989. Kopalne szczątki zwierzęce. In Jahn, A., Kozłowski, S., and Wiszniowska, T. (eds.), Jaskinia Niedźwiedzia w Kletnie – Badania i udostępnianie, p. 255-279. Zakład Narodowy Ossolińskich, Wydawnictwo Polskiej Akademii Nauk, Wrocław - Warszawa - Kraków - Gdańsk – Łódź.
Wiszniowska, T., Bieroński, J., and Pakiet, M. 1996. Paleoekologia Masywu Śnieźnika, p. 47 – 5. In Jahn, A., Kozłowski, S., and Pulina, M. (eds.), Masyw Śnieźnika – zmiany w środowisku przyrodniczym. Polska Agencja Ekologiczna SA, Wydawnictwa PAE, Warszawa.
