

 Are there two distinct types of hypocone in Eocene primates? The 'pseudohypocone' of notharctines revisited
Are there two distinct types of hypocone in Eocene primates? The 'pseudohypocone' of notharctines revisited
Article number: 15.3.26A
https://doi.org/10.26879/329
Copyright Society for Vertebrate Paleontology, October 2012
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 22 May 2012. Acceptance: 20 September 2012
{flike id=306}
ABSTRACT
Upper molars of modern humans and most extant primates have four cusps that have evolved from the original tribosphenic tooth of therian mammals. These include the three cusps of the original trigon (e.g., paracone, metacone, and protocone), and the addition of the distolingual cusp or hypocone. Among Eocene primates of the family Adapidae, a distinction has long been made between a "true" hypocone associated with the lingual cingulum (adapine form) and a "pseudohypocone" associated with the distal margin of the protocone (notharctine form). The developmental processes underlying these two types of distolingual cusp are unknown, and the validity of the distinction is based on phylogenetic utility and homology rather than cusp position, as in other mammalian groups. To address this issue we use micro-computed tomography to reveal the morphology of the hypocone and associated cusps and crests on the enamel-dentine junction (EDJ). The EDJ preserves the initial steps of tooth crown development and can be used to clarify detailed aspects of crown morphology in variably worn or damaged fossil teeth. Our study sample includes both adapine species from Europe and notharctines from North America. We confirm that the pseudohypocone found among notharctines is a true cusp since it forms as a dentine horn during crown development. Our results also confirm that these two forms of hypocone are developmentally distinct and have evolved convergently in these two primate clades. A review of the paleontological literature suggests that, in spite of the fact that homoplasy is rampant among mammalian clades with respect to the development of the hypocone, only among the notharctines do we find an alternative name for this cusp.
Robert L. Anemone. Department of Anthropology, Western Michigan University, Kalamazoo, MI, USA 49008 anemone@wmich.edu
Matthew M. Skinner. Department of Anthropology, University College London, London, United Kingdom and Department of Human Evolution, Max Planck Institute for Evolutionary Anthropology, Leipzig, Germany m.skinner@ucl.ac.uk
Wendy Dirks. School of Dental Sciences, Newcastle University, Newcastle upon Tyne, United Kingdom wendy.dirks@newcastle.ac.uk
Key Words: odontogenesis; primates; adapids; crown morphology; dentition; hypocone
Final Citation: Anemone, Robert L., Skinner, Matthew M., and Dirks, Wendy. 2012. Are there two distinct types of hypocone in Eocene primates? The ‘pseudohypocone’ of notharctines revisited. Palaeontologia Electronica Vol. 15, Issue 3;26A,13p;
palaeo-electronica.org/content/2012-issue-3-articles/306-hypocones-in-eocene-adapids
INTRODUCTION
Teeth tend to be the most commonly preserved material in most fossil mammal assemblages, and the study of dental crown morphology can yield a wealth of useful information concerning the diet, behavior, and phylogenetic relationships among living and fossil mammals, including Primates. While morphological features of the occlusal surface of primate teeth comprise the primary dataset for understanding phylogenetic relations among fossil taxa (Rose, 2006), interpretations of these data are often constrained by a lack of information on the developmental processes that result in features of dental morphology like cusps, crests, and cingula. Specifically, determining whether features can be considered homologous for the purpose of phylogenetic reconstruction, as well as determining the polarity of homologous character states, can be informed by a detailed understanding of the underlying developmental processes underlying trait form and variation (Jernvall et al., 2008). While some consider it necessary to have a molecular-based phylogeny in order to identify whether traits are homologous or homoplastic (Wake et al., 2011), gross morphology and ontogenetic development can also yield important information (particularly in fossil taxa) for determining the phylogenetic valence of particular structures
One long-standing controversy in primate evolution with respect to molar occlusal morphology and its phylogenetic significance concerns the identity and origin of the distolingual cusp among upper molars in Eocene adapid primates. Stehlin (1916) was the first to notice that the hypocones of North American notharctines were different enough from the closely related European adapines to suggest an independent (i.e., convergent) origin of this cusp in the two clades. Gregory (1922) concurred and clearly described and illustrated the morphological differences between what Stehlin had called the true hypocone of adapines and the pseudohypocone of notharctines. Gregory suggested that true hypocones develop from the lingual cingulum (or postero-internal cingulum) in adapines and in most other mammals, while pseudohypocones arise in notharctines "as a budding or outgrowth from the posterior slope of the protocones" (Gregory, 1922, p. 130). This latter structure is known variously in the literature as the postprotocrista, postprotocone fold, or Nannopithex fold, and it is typically described as a crest running disto-lingually from the protocone. In his revision of the plesiadapiform family Phenacolemuridae, Simpson (1955) questioned the morphological distinction between hypocones and pseudohypocones among Eocene primates and hinted at the difficulties in determining homologies between cusps on the teeth of fossil primates when he stated (Simpson, 1955, p. 435):
"I strongly question, however, whether independent origin of the cusp from, say, a cingulum in two different groups makes it in an objective sense any more the "same" cusp than if it arose in the same place but not from a cingulum...Whether the cusp originates from the "Nannopithex-fold" and is a "pseudohypocone" or from the cingulum and is a "hypocone" seems to me a distinction without a difference."
Butler (1956, 1963) also doubted whether hypocones could or should be distinguished among adapids based on his sense of the large range of variation in this cusp at several different hierarchical levels, including among different mammalian taxa and within individual taxa, and even within a single individual. Speaking of the importance of taking metameric variation into account when interpreting crown morphology Butler (1963, p. 12) states:
"When a fourth cusp arises it stands on the same lingual marginal ridge as the protocone, and thus it has the appearance of a pseudohypocone. It may retain this appearance throughout the series, or alternatively it may lose its connection with the protocone on the more distal teeth, and then it stands alone, as if it were part of a cingulum. The pseudohypocone and the true hypocone therefore intergrade, and there is no point in using the term pseudohypocone at all."
Returning to the question of the evolution of the hypocone later in his long career, Butler (2000) reversed his position and supported the distinction between hypocones and pseudohypocones among Eocene adapids on essentially the same terms as described earlier by Gregory (1922).
In his discussion of a number of European Eocene forms, Simons (1962, p. 7) recognized the presence of the Nannopithex-fold in the type specimen of Cantius eppsi from Abbey Wood, but noted "no indication of an incipient hypocone element, other than a slight thickening of the posterolingual part of the basal protocone cingulum." To further complicate matters, Simons (1962) asserted that both a pseudohypocone and true hypocone can be found in a single individual (AMNH 15022) of the North American notharctine Pelycodus (= Cantius) (Gingerich and Haskin, 1981) and of the European adapine Caenopithecus (Eh. 727). This suggested to him that "a latency for producing pseudohypocones, as in the line leading to Notharctus as well as for true hypocone production in Adapis, probably existed in the ancestral adapid-notharctid stock" (Simons, 1962, p. 30). While we have not had the opportunity to examine the specimen of Caenopithecus in the collections at Basel, we interpret AMNH 15022 as having a typical notharctine pseudohypocone arising from the Nannopithex fold with a typical lingual cingulum, and remain skeptical of the claim for both types of hypocones being present in single individuals of Eocene adapids.
The hypocone – pseudohypocone distinction has continued to be a matter of contention in primate systematics both within the adapiforms and in studies of anthropoid origins. In a discussion of adapiform systematics and phylogeny, Godinot (1998:230) says, "One might even suspect that Smilodectes' hypocone is not homologous with that of other notharctines: its position is intermediate between a cingular hypocone and the pseudohypocone typical of other notharctines." In their critical review of the Asian origins for Anthropoidea, Ciochon and Holroyd (1994, p. 146) state, "The presence or absence of a pseudohypocone and/or a hypocone in Pondaungia is critical to understanding its phylogenetic affinities," and suggest that the disto-lingual cusp in Pondaungia is a pseudohypocone. They continue:
"...(T)he degree of separation of protocone and hypocone is not the relevant attribute in recognizing the presence of a pseudohypocone. Among primates, pseudohypocones occur uniquely in notharctine adapids and are recognized by their place of origin, independent of their degree of separation from the protocone... The difference in terminology reflects the fact that the distolingual cusp in northactines arises from the postprotocrista rather than from the distolingual cingulum (a 'true' hypocone)" (Ciochon and Holroyd, 1994, p. 146).
Shigehara et al. (2002) dispute this interpretation of the distolingual cusp in Pondaungia in a lengthy discussion of what constitutes a pseudohypocone. Citing Gregory (1920), they argue that a pseudohypocone requires twinning of a cusp located on the Nannopithex fold with the protocone. They also note that "...(M)any primates have hypocones that are not attached to the distolingual cingulum, for example Callicebus, so the absence of a connection between the distolingual cusp and the distolingual cingulum is not a reliable guide to the identification of a pseudohypocone" (Shigehara et al., 2002, p. 154).
In his major revision of New World monkeys, Hershkovitz (1977) revisited the issue of the nature and types of hypocones and offered the most cogent modern rejection of the idea that hypocones and pseudohypocones are distinct embryological or morphological features of Eocene primates. Following Remane's (1960) suggestion that primate hypocones always arise from the lingual cingulum, Hershkovitz argued that Gregory, Stehlin and most other previous authors had misidentified the dental structures associated with the presence of a pseudohypocone among North American notharctines. Rather than being the primary lingual cingulum from which hypocones develop, Hershkovitz identified the cingulum found alongside pseudohypocones in notharctines as a secondary cingular structure. Hershkovitz (1977, p. 293) concluded that his "studies of large series of mammalian molars and premolars of all geologic ages and stages of development revealed no evidence of the origin of a hypocone or 'pseudohypocone' elsewhere than from the primary posterolingual cingulum."
In summary, debates continue concerning the legitimacy of a distinction between the type of hypocone seen in adapines and that seen in notharctines, and at the center of this debate lie developmental issues concerning the presence of primary and secondary cingula and their spatial relationships to the primary cusps, including the hypocone. In our opinion, the terminology that we use to describe these morphological features is critically important because of its influence on how we think about their phylogenetic significance. Several critical questions are raised by our review of the literature on the hypocone among Eocene Adapidae and these questions form the raison d'etre of this paper:
1. Can we better understand the true nature of the distinction between hypocones and pseudohypocones by using 3-dimensional imaging to explore the morphology and developmental origins of the distolingual cusp?
2. Do the morphological and/or developmental differences between the hypocone and pseudohypocone warrant the use of two different names for the distolingual cusp in Eocene adapids?
The Importance of the EDJ
Recent research has demonstrated that examination of the internal structure of teeth, and in particular the surface of the dentine crown, can clarify the developmental processes underlying aspects of tooth crown morphology and facilitate comparisons between different taxa (Ortiz et al., 2012; Bailey et al., 2011; Skinner and Gunz, 2010; Skinner et al., 2008, 2009, 2010). This is because the surface of the dentine crown, often referred to as the enamel-dentine junction (or EDJ), preserves the morphology of the basement membrane of the developing tooth germ prior to mineralization (Butler, 1956; Nager, 1960; Krause and Jordan, 1965). Thus, the EDJ can be thought to preserve the first stage of crown development in which the cusps and crests appear. Subsequently, enamel tissue is deposited over the EDJ to yield the final form of the unworn tooth crown. While there is some overlap in these two processes that involves growth of the basement membrane between the initiation of mineralization at cusp tips and the time of coalescence of the mineralizing front that terminates changes in EDJ shape, it is heuristic to consider these processes separately. Furthermore, since the original shape of the tooth crown is often not preserved in fossils due to either wear or destructive taphonomic processes, the EDJ can yield important information about the original shape of the tooth crown, the presence of cusps, and the early stage of tooth development.
This study uses microtomography to assess the morphology of the EDJ in a sample of adapines and notharctines in order to shed light on the developmental relationships among cusps and crests associated with the hypocone. Specifically, we address whether there is evidence to support the hypothesis that the hypocone in these groups forms as the result of fundamentally distinct developmental processes, and are thus homoplastic features. Finally, we reevaluate the utility of distinguishing between true hypocones and pseudohypocones among primates, and set this discussion within a wider phylogenetic context by surveying the literature concerning the diversity of hypocones among living and fossil mammals.
MATERIALS AND METHODS
The study sample is listed in Table 1 and consists of maxillary dentitions of the following species: Adapisparisiensis, Leptadapisleenhardti, Notharctusventicolus, N. nunienus, and Cantiusabditus. Each specimen was microCT scanned using either a Skyscan 1172 scanner or an Actis BIR 225 scanner. The resolution of the resultant reconstructed image stacks ranged from 10-30 microns. Each image stack was filtered using a median filter and mean-of-least-variance filter to clarify tissues boundaries. Enamel and dentine tissues were segmented using a semi-automatic segmentation protocol. Digital surface models (.ply format) were generated from the segmentations using the surface generation module in Avizo 6.3 (with unconstrained smoothing). Manual editing of surface models to fill holes or repair cracks was done in Geomagic Studio 11. A number of specimens exhibited similar density differences in enamel and dentine preventing segmentation of the tissues and examination of the EDJ (see Table 1). In these cases, observations were limited to the outer enamel surface morphology.
RESULTS
The Hypocone in European Adapinae
 Surface models of the outer enamel surface and EDJ (when available) of two European adapines are illustrated in Figure 1. The close association between the hypocone and the lingual cingulum is evident in all specimens. A clear dentine horn for the hypocone can be seen on the first and second molar of YPM-PU 11499 (Figure 1.4). This dentine horn is similar in its morphology to adjacent cusps and to dentine horns seen across primates. Of particular relevance to this study is the lack of morphological association between the dentine horn for the hypocone and the protocone. The post-protocone crest runs directly to the metacone, and there is no evidence of a postprotocrista or Nannopithex fold. The Leptadapis specimen (YMP-PU 11481, Figure 1.5-1.6) exhibits similar hypocone morphology to the Adapis specimens (Figure 1.1-1.4) with regard to the presence and positioning of a hypocone dentine horn and the absence of a postprotocone fold. It is worth noting the diminishing size of the hypocone from the first to the third molar in YPM-PU 11481, and the third molar in particular is a good example of minor expression of this cusp in adapines.
Surface models of the outer enamel surface and EDJ (when available) of two European adapines are illustrated in Figure 1. The close association between the hypocone and the lingual cingulum is evident in all specimens. A clear dentine horn for the hypocone can be seen on the first and second molar of YPM-PU 11499 (Figure 1.4). This dentine horn is similar in its morphology to adjacent cusps and to dentine horns seen across primates. Of particular relevance to this study is the lack of morphological association between the dentine horn for the hypocone and the protocone. The post-protocone crest runs directly to the metacone, and there is no evidence of a postprotocrista or Nannopithex fold. The Leptadapis specimen (YMP-PU 11481, Figure 1.5-1.6) exhibits similar hypocone morphology to the Adapis specimens (Figure 1.1-1.4) with regard to the presence and positioning of a hypocone dentine horn and the absence of a postprotocone fold. It is worth noting the diminishing size of the hypocone from the first to the third molar in YPM-PU 11481, and the third molar in particular is a good example of minor expression of this cusp in adapines.
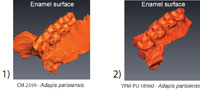
The Pseudohypocone in North American Notharctinae
 Figure 2 illustrates enamel and EDJ expression of the pseudohypocone of the Notharctus specimens in our sample. The cusp-like morphology of the pseudohypocone as seen at the enamel surface would suggest an underlying dentine horn (Figure 2.1-2.3). The presence of an obvious dentine horn underlying the distolingual cusp in both the first and second molars of CM 43370 (Figure 2.4) clearly defines this as a true cusp, regardless of the fact that it derives from the postprotocone fold rather than from the lingual cingulum as in Adapis and Leptadapis. In addition, the postprotocone fold seems to be homologous to a similar crest that runs distally from the protocone in the upper fourth premolars of CM 22062 and CM 43370 (Figure 2.1-2.3). Figure 3 illustrates enamel and EDJ expression of the pseudohypocone in Cantius. In the majority of specimens the pattern seen in Cantius is similar to that seen in Notharctus, but not all specimens have a distolingual cusp. Many of the isolated teeth in our study sample exhibit only a postprotocrista or Nannopithex fold running distally from the protocone (Figure 3.4), but with no enamel or dentine horn for a distolingual cusp. This is entirely predictable in light of the well-established variability of pseudohypocone development among Cantius species from the early Eocene (Gingerich and Haskin, 1981; Gingerich and Simons, 1977).
Figure 2 illustrates enamel and EDJ expression of the pseudohypocone of the Notharctus specimens in our sample. The cusp-like morphology of the pseudohypocone as seen at the enamel surface would suggest an underlying dentine horn (Figure 2.1-2.3). The presence of an obvious dentine horn underlying the distolingual cusp in both the first and second molars of CM 43370 (Figure 2.4) clearly defines this as a true cusp, regardless of the fact that it derives from the postprotocone fold rather than from the lingual cingulum as in Adapis and Leptadapis. In addition, the postprotocone fold seems to be homologous to a similar crest that runs distally from the protocone in the upper fourth premolars of CM 22062 and CM 43370 (Figure 2.1-2.3). Figure 3 illustrates enamel and EDJ expression of the pseudohypocone in Cantius. In the majority of specimens the pattern seen in Cantius is similar to that seen in Notharctus, but not all specimens have a distolingual cusp. Many of the isolated teeth in our study sample exhibit only a postprotocrista or Nannopithex fold running distally from the protocone (Figure 3.4), but with no enamel or dentine horn for a distolingual cusp. This is entirely predictable in light of the well-established variability of pseudohypocone development among Cantius species from the early Eocene (Gingerich and Haskin, 1981; Gingerich and Simons, 1977).
 One specimen that exhibits unique morphology in our sample is WMU 2588 (Figure 3.5-3.8). The reduced distolingual crown margin of this specimen suggests that it is a third molar, and would thus be expected to lack a hypocone. What is unique about this specimen is the presence of two dentine horns on the crest that runs distobucally from the protocone. In this specimen the intermediate dentine horn could be interpreted as a pseudohypocone, however, it is located on the crest running towards the metacone rather than directly distal from the protocone. The adjacent and more diminutive dentine horn could be homologous with the metaconule. Interpreting the significance of this kind of variation under our current understanding of the genetic processes underlying cusp patterning within the primate clade is discussed below.
One specimen that exhibits unique morphology in our sample is WMU 2588 (Figure 3.5-3.8). The reduced distolingual crown margin of this specimen suggests that it is a third molar, and would thus be expected to lack a hypocone. What is unique about this specimen is the presence of two dentine horns on the crest that runs distobucally from the protocone. In this specimen the intermediate dentine horn could be interpreted as a pseudohypocone, however, it is located on the crest running towards the metacone rather than directly distal from the protocone. The adjacent and more diminutive dentine horn could be homologous with the metaconule. Interpreting the significance of this kind of variation under our current understanding of the genetic processes underlying cusp patterning within the primate clade is discussed below.
DISCUSSION
Analysis of the expression of the distolingual cusp on the enamel surface of molars of adapines and notharctines is consistent with the two distinct patterns that have been well-known to paleontologists since the early 20th century. The use of micro CT scanning, however, allows us to document for the first time the underlying morphology of the EDJ in these fossil taxa. This examination informs our consideration of the distinction between true and pseudohypocones in primate evolution by providing critical developmental evidence concerning the ontogeny of these cusps. Figure 4 illustrates an upper second molar of Adapis (YPM-PU 11499) and Notharctus (CM 43370) as well as a schematic illustration that labels relevant features following Hershkovitz (1977; and in particular his figure V.14 on p. 288). Animated pdfs of these two teeth can be found in the Appendix. Our results indicate that Stehlin (1916), Gregory (1922) and other early students of primate evolution who suggested an important distinction between the form of the distolingual cusp in Eocene adapids were correct. The distolingual cusp of adapines is derived from the lingual cingulum, while the distolingual cusp among notharctines is derived from the postprotocone or Nannopithex fold. We suggest that the presence of a dentine horn underlying this cusp in both groups definitively marks the distolingual cusp in all Eocene adapids as a true cusp, but that these two cusps are clearly homoplastic structures. Furthermore, we suggest that a coherent picture of the evolution of the hypocone and related dental structures among primates can be illuminated by a consideration of our results in conjunction with the model presented by Hershkovitz (1977).
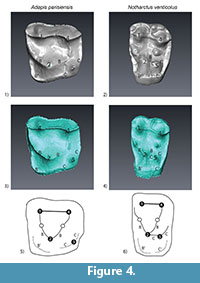 In Hershkovitz's (1977) model, the postprotocone fold and the "pseudohypocone" which arises from it in notharctines is homologous with what he calls the primary posterolingual cingulum in adapines, from which arises the "true hypocone". In our Figure 4, this cingulum is labeled "C". If this interpretation is correct, the "pseudohypocone" and "true hypocone" in Eocene adapids are homologous structures, and the difference between adapines and notharctines is that 1) no secondary posterolingual cingulum (C') has formed in Adapis and 2) there is no crest linking the primary anterolingual cingulum (B) and the primary posterlingual cingulum (C) in adapines. A potential problem with Hershkovitz's interpretation, in our opinion, is that the cingulum in Adapis is necessarily composed of a secondary cingulum anteriorly (B') and a primary cingulum posteriorly (C). We disagree with Butler (1963) that these two forms intergrade with one another, and we are not convinced that homologizing the Nannopithex fold with part of the primary posterolingual cingulum is the correct interpretation. An important question is: are the morphological differences in the distolingual cusp and associated crests and cingula significant enough to indicate two fundamentally different developmental processes? While our results and interpretation would benefit from the inclusion of a wider variety of fossil Eocene primates, we conclude that the developmental evidence presented here strongly suggests that the hypocone and pseudohypocone have evolved convergently in adapines and notharctines, respectively.
In Hershkovitz's (1977) model, the postprotocone fold and the "pseudohypocone" which arises from it in notharctines is homologous with what he calls the primary posterolingual cingulum in adapines, from which arises the "true hypocone". In our Figure 4, this cingulum is labeled "C". If this interpretation is correct, the "pseudohypocone" and "true hypocone" in Eocene adapids are homologous structures, and the difference between adapines and notharctines is that 1) no secondary posterolingual cingulum (C') has formed in Adapis and 2) there is no crest linking the primary anterolingual cingulum (B) and the primary posterlingual cingulum (C) in adapines. A potential problem with Hershkovitz's interpretation, in our opinion, is that the cingulum in Adapis is necessarily composed of a secondary cingulum anteriorly (B') and a primary cingulum posteriorly (C). We disagree with Butler (1963) that these two forms intergrade with one another, and we are not convinced that homologizing the Nannopithex fold with part of the primary posterolingual cingulum is the correct interpretation. An important question is: are the morphological differences in the distolingual cusp and associated crests and cingula significant enough to indicate two fundamentally different developmental processes? While our results and interpretation would benefit from the inclusion of a wider variety of fossil Eocene primates, we conclude that the developmental evidence presented here strongly suggests that the hypocone and pseudohypocone have evolved convergently in adapines and notharctines, respectively.
The results of this paper highlight the importance of understanding the developmental processes underlying tooth crown morphology in order to properly characterize and interpret its variation within and among species. Defining whether or not a feature is a true cusp is facilitated by the ability to identify a dentine horn at the EDJ. The dentine horn can be considered as the morphological remnant of the presence of a secondary enamel knot during the development of the tooth crown. We suggest that it is difficult to consider a feature as a cusp in the absence of some evidence that it arose from an enamel knot. The metameric variation in pseudohypocone morphology in the Notharctus specimen (Figure 2.4) highlights the fact that determination of a feature as a cusp may require finding specimens that exhibit marked expression of the feature. This kind of variation is suggestive of a scenario in which it is the interaction between various aspects of tooth crown morphogenesis that dictates whether, and to what degree, cusps appear.
Cusp development is considered to be an iterative process involving the sequential appearance of secondary enamel knots on the developing tooth crown (Jernvall and Jung, 2000). The pattern of dentine horns seen on the distal margin of WMU 2588 is consistent with this process and resembles very closely the patterning of accessory dentine horns seen on the distal margin of chimpanzee lower molars (Skinner and Gunz, 2010). Specimens such as WMU 2588 highlight the fact that this process of cusp patterning can be influenced by other aspects of tooth crown development. In this case it is likely that the overall size of the tooth crown (being a reduced and somewhat mesiodistally compressed third molar) and the relationship between the trigon crest and the distal margin of the tooth influenced the patterning of dentine horns on the crown.
Simons (1962) has suggested that Pelycodus (=Cantius) frugivorus, a notharctine primate, exhibits a morphological pattern that has the potential to be ancestral to both adapines and notharctines. This is due to the proposed presence of both a pseudohypocone (in the notharctine sense) and an incipient hypocone on the distolingual margin of the cingulum (see specimen AMNH 15022, Plate XXXV, Gregory, 1920). The results of our study suggest that this hypothesis should be re-evaluated. Recognizing that our sample is small, we found no clear evidence in our notharctine sample of a dentine horn on the cingulum that would be consistent with a hypocone. In addition, our examination of several hundred specimens from multiple species of Cantius from our collections in the Great Divide Basin has never revealed a specimen with both a true and a pseudohypocone. We recommend that attempts be made to identify specimens in museum collections whose enamel morphology is suggestive of this configuration and examine the underlying EDJ. From a developmental perspective it would be a significant finding that a species possesses essentially two types of hypocone on the same crown and thus clearly evidence of this fact should be sought out.
Butler believed that variation in hypocone development may largely be attributed to variations in the timing of appearance of the lingual cusps during ontogeny. This begs the question as to whether the difference seen between adapines and notharctines is indeed just a difference in timing or something more fundamental. If it is consistent within a species and the metameric variation does not suggest that two morphs 'overlap', then we believe they can be considered distinct and for particular taxonomic comparisons will carry systematic valence. Butler (1956, p. 47) noted that new "cusps may also arise from growth zones in the valleys between the cusps, which may be regarded as fragments of the zona cingularis." Thus, what might be important is the distinction of whether a cusp forms on a cingulum, or in association with another cusp; that is, externally or internally on the crown. In the case of Eocene notharctines, the "pseudohypocone" does not form from a cingulum.
CONCLUSION
When one considers the larger picture of the evolution of mammalian molar occlusal patterns, and in particular the addition of a hypocone to the primitive tribosphenic molar, it seems that homoplasy is the rule rather than the exception. Hunter and Jernvall (1995) demonstrated that the hypocone has evolved independently more than twenty times among mammals, often as in adapines as an outgrowth of the posterolingual cingulum, but also as an inflated metaconule (as in deer and other artiodactyls) (Black, 1978; Hunter and Jernvall, 1995). They suggest that clades, which have evolved a hypocone, tend to have higher species diversity and to typically be herbivorous, and they conclude that the hypocone fits Mayr's (1963) definition of a "key innovation" since it has led to the successful invasion of new, herbivorous adaptive zones for many different mammal clades. Thus, the convergent acquisition of a hypocone among two closely related subfamilies of Eocene primates should be considered the norm among mammals, and no necessary cause for the creation of new cusp nomenclature (i.e., pseudohypocone). Ungar (2010) has argued the same, suggesting that "nomenclatural pandemonium" has resulted from updating the naming system developed from Cope and Osborn's original misinterpretation of cusp homologies, stating, "The least confusing solution... is to continue to use Osborn's basic terminology, acknowledging that cusp names no longer imply serial homology." (Ungar, 2010:13)
The presence of a dentine horn beneath the pseudohypocone of notharctines suggests the term itself is a misnomer. The distolingual cusp in notharctines is neither a "pseudo" cusp (since like all true cusps, it originates with a dentine horn), nor is it a "pseudo" hypocone (because its position as the distolingual cusp on the quadrate upper molar defines it as a hypocone). Specifically, in minor forms of expression the feature is a crest running distally from the protocone. In marked forms of expression (illustrated by the first and second molars of the Notharctus specimen CM 43370) a clear dentine horn develops. The distinction between a 'strong Nannopithex fold' and a pseudohypocone likely reflects the variation documented in this study between different molars within the same tooth row. Thus, this feature should be considered as a hypocone that has evolved convergently, with respect to adapines, through the appearance of a distal crest and associated dentine horn distal to the protocone. Our examination of the EDJ confirms previous observations, based on examination of the enamel surface, that the hypocone of Adapis develops as a true cusp (i.e., deriving from a dentine horn) that arises from the cingulum. The utility of the EDJ for clarifying the 'cusp' status of crown features encourages re-assessment of Simon's suggestion that specimens of P. frugivorus show both a pseudohypocone and an incipient hypocone on the cingulum.
ACKNOWLEDGMENTS
This research was supported by Jean-Jacques Hublin, Department of Human Evolution, Max Planck Institute for Evolutionary Anthropology and the Max Planck Society. For access to specimens in their care, we thank E. Sargis and D. Brinkman of the Yale Peabody Museum of Natural History, C. Beard and A. Tabrum of the Carnegie Museum of Natural History, and T. Koppe and the Greifswald Anatomy Museum. We also thank P. Schoenfeld for CT scanning assistance. This paper was substantially improved as a result of the recommendations of two anonymous reviewers.
REFERENCES
Bailey, S., Skinner, M., and Hublin, J-J. 2011. What lies beneath? An evaluation of lower molar trigonid crest patterns based on both dentine and enamel expression. American Journal of Physical Anthropology, 145:505-518.
Black, C. 1978. Paleontology and geology of the Badwater Creek Area, Central Wyoming. Part 14. The Artiodactyls. Annals of Carnegie Museum, 47:223-259.
Butler, P. 1956. The ontogeny of molar pattern. Biological Reviews, 31:30-70.
Butler, P. 1963. Tooth morphology and primate evolution, p. 1-13. In Brothwell, D.R., editor. Dental Anthropology. New York, Pergamon Press.
Butler, P. 2000. The evolution of tooth shape and tooth function in primates, p 201-211. In Teaford M., Smith, M., and Ferguson, M., (eds.), Development, Function and Evolution of Teeth. Cambridge: Cambridge University Press.
Ciochon, R., and Holroyd, P. 1994. The Asian origin of Anthropoidea revisited, p. 143-162. In Fleagle, J. and Kay, R., (eds.), Anthropoid Origins. New York: Plenum Press.
Gingerich, P.D. and Haskin, R.A. 1981. Dentition of early Eocene Pelycodus jarrovii (Mammalia, Primates) and the generic attribution of species formerly referred to Pelycodus. Contributions from the Museum of Paleontology, University of Michigan, 25(17):327-337.
Gingerich. P,D, and Simons, E.L. 1977. Systematics, phylogeny, and evolution of early Eocene Adapidae (Mammalia, Primates) in North America. Contributions from the Museum of Paleontology, University of Michigan, 24(22):245-279.
Godinot, M. 1998. A summary of Adapiform systematics and phylogeny. Folia Primatologica, 69:218-249.
Gregory, W. 1922. The Origin and Evolution of the Human Dentition. Williams and Wilkins, Baltimore.
Gregory, W.K. 1920. On the structure and relations of Notharctus, an American Eocene primate. Memoirs of the American Museum of Natural History, 3:51-243.
Hershkovitz, P. 1977. Living New World Monkeys (Platyrrhini). With an Introduction to Primates, Vol. 1. Chicago: University of Chicago Press.
Hunter, J. and Jernvall, J. 1995. The hypocone as a key innovation in mammalian evolution. Proceedings of the National Academy of Sciences, USA, 92:10718-10722.
Jernvall, J. and Jung, H. 2000. Genotype, phenotype, and developmental biology of molar tooth characters. Yearbook of Physical Anthropology, 43:171-190.
Jernvall, J., Gilbert, C., and Wright, P. 2008. Peculiar tooth homologies of the greater bamboo lemur (Prolemur = Hapalemur simus). When is a paracone not a paracone? p. 335-342. In Fleagle J. and Gilbert, C, (eds.), Elwyn Simons: A Search for Origins. New York: Springer.
Krause, B. and Jordan, R. 1965. The Human Dentition before Birth. Lea and Febiger, Philadelphia.
Mayr, E. 1963. Animal Species and Evolution. Cambridge, MA: Harvard University Press.
Nager, G. 1960. Der vergleich zwischen dem raumlichen verhalten des dentinkronenreliefs und dem schmelzrelief der zahnkrone. Acta Anatomica, 42:226-250.
Ortiz, A., Skinner, M.M., Bailey, S.E., Hublin, J.-J. 2012. Carabelli’s trait revisted: an examination of mesiolingual features at the enamel-dentine junction and enamel surface of Pan and Homo sapiens upper molars. Journal of Human Evolution 63: 586-596.
Remane, A. 1960. Zahne und Gebis, p. 637-846. In Hofer, H., Schultz, A., and Starck, D. (eds.), Primatologie. Basel: S. Karger.
Rose, K. 2006. The Beginning of the Age of Mammals. The Johns Hopkins University Press, Baltimore.
Shigehara, N., Takai, M., Kay, R., Aung, A., Soe, A., Tun, S., Tsubamoto, T., and Thein, T. 2002. The upper dentition and face of Pondaungia cotteri from central Myanmar. Journal of Human Evolution, 43:143-166.
Simons, E.L. 1962. A new Eocene primate genus, Cantius, and a revision of some allied European lemuroids. Bulletin of the British Museum of Natural History, Geologt, 7:1-36.
Simpson, G.G. 1955. The Phenacolemuridae, new family of early Primates. Bulletin of the American Museum of Natural History, 105(5):411-442.
Skinner, M. and Gunz, P. 2010. The presence of accessory cusps in chimpanzee lower molars is consistent with a patterning cascade model of development. Journal of Anatomy, 217:245-253.
Skinner, M.M., Evans, A., Smith, T., Jernvall, J., Tafforeau, P., Kupczik, K., Olejniczak, A.J., Rosas, A., Radovčić, J., Thackeray, J.F., Toussaint, M., Hublin, J.-J. 2010. Brief communication: Contributions of enamel-dentine junction shape and enamel deposition to primate molar crown complexity. American Journal of Physical Anthropology 142: 157-163.
Skinner, M, Wood, B., and Hublin, J-J. 2009. Protostylid expression at the outer enamel surface and at the enamel-dentine junction of lower molars in extant and fossil hominoids. Journal of Human Evolution, 56:76-85.
Skinner, M., Wood, B., Boesch, C., Olejniczak, A., Rosas, A., Smith, T., and Hublin, J-J. 2008. Dental trait expression at the enamel-dentine junction of lower molars in extant and fossil hominoids. Journal of Human Evolution, 54:173-186.
Skinner, M,, Wood, B., and Hublin, J-J. 2009. Protostylid expression at the outer enamel surface and at the enamel-dentine junction of lower molars in extant and fossil hominoids. Journal of Human Evolution, 56:76-85.
Stehlin, H.G. 1916. Die Säugetiere des schweizerischen Eocaens: Critisches Catalog der Materialen Siebenter Teil, Zweiter Hafte. Abh Schweiz Paläont Gesellsch, 41:1299-1552.
Ungar, P.S. 2010. Mammal Teeth: Origin, Evolution, and Diversity. Baltimore, MD: Johns Hopkins University Press.
Wake, D.B., Wake, M.H., and Specht, C.D. 2011. Homoplasy: from detecting pattern to determining process and mechanism of evolution. Science, 331(6020):1032-1035.
