

 Motion range of the manus of Plateosaurus engelhardti von Meyer, 1837
Motion range of the manus of Plateosaurus engelhardti von Meyer, 1837
Article number: 17.1.12A
https://doi.org/10.26879/428
Copyright Palaeontological Association, March 2014
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 19 August 2013. Acceptance: 1 March 2014
{flike id=692}
ABSTRACT
The mode of locomotion of the basal sauropodomorph Plateosaurus engelhardti, known from numerous finds from the late Triassic of Central Europe, has been extensively debated. Some early and recent research results indicate that the forelimb could not play role in quadrupedal locomotion. Other authors suggested facultative or even permanent quadrupedality. This would require adaptations of the range of motion and the stability of the manual digits to the high forces caused by locomotion. An analysis of the hyperextension capabilities of the hand can therefore determine if the manus is adapted for locomotion. This study examines the capabilities of the manus of P. engelhardti using digital 3D modeling. The motion ranges of the digits were simulated in a computer-aided engineering (CAE) program, and the hyperextension capability of the entire manus was tested.
We find that the hand of Plateosaurus was not able to support the animal during quadrupedal locomotion, but may rather have been a specialized grasping organ. Therefore, P. engelhardti must have been an obligate biped.
Stefan Reiss. Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Rheinische Friedrich-Wilhelms-Universität Bonn, Germany. general-rabe@gmx.de
Heinrich Mallison. Museum für Naturkunde - Leibniz-Institut für Evolutions - und Biodiversitätsforschung, Berlin, Germany. heinrich.mallison@mfn-berlin.de
Keywords: Plateosaurus; range of motion; virtual palaeontology
Final citation: Reiss, Stefan and Mallison, Heinrich. 2014. Motion range of the manus of Plateosaurus engelhardti von Meyer, 1837. Palaeontologia Electronica Vol. 17, Issue 1;12A; 19p. https://doi.org/10.26879/428
palaeo-electronica.org/content/2014/692-plateo-hand
INTRODUCTION
The osteology of Plateosaurus engelhardti von Meyer, 1837, a basal sauropodomorph, was first described in detail by von Huene (1926). Plateosaurus is one of the best known dinosaurs with over 100 individuals found at numerous sites in Central Europe, including the bone-beds at Frick (Switzerland), Halberstadt, and Trossingen (both Germany) (Sander, 1992; Moser, 2003; Klein, 2004; Mallison, 2010a, 2010b). There is no clear consensus on the species taxonomy. It seems likely that all individuals with cranial material from Trossingen, Frick and Halberstadt belong to one species, P. erlenbergensis (Galton, 2001; Prieto-Márques and Norell, 2011). A detailed study of the sacra found P. engelhardti to be valid and the sacrum of the type series sufficiently rich in characters that it was designated as lectotype (Moser, 2003). Moser also concluded that P. erlenbergensis von Huene, 1905 and its junior synonym P. longiceps Jaekel, 1913, to which the Halberstadt and Trossingen material was previously referred Galton (2001), are junior synonyms of P. engelhardti. We here follow Moser (2003) in regarding all diagnostic material from the upper Löwenstein Formation and the Trossingen Formation as P. engelhardti, because no study conducted after Moser's 2003 review (e.g., Prieto-Marques and Norell, 2011, who refer the Trossingen material to P. erlenbergensis) has addressed the issue of sacral characters, and thus made a convincing case for the presence of a different species in Trossingen or Halberstadt. According to von Huene (1926) and Yates (2003a), Sellosaurus is a junior synonym of Plateosaurus, but the older species P. (Sellosaurus) gracilis is not the focus of this study. The great similarity of the two species, especially in the forelimb, likely means that conclusion on P. engelhardti also apply to P. gracilis.
Based on the dating of the strata in which specimens were found, Plateosaurus engelhardti apparently occurred in the late Norian and disappeared in the Rhaetian (Galton, 2001; Yates, 2003a). Plateosaurus engelhardti reached an estimated maximum size between a total length of 7.5 m (Klein, 2004) and 10 m, showing significant developmental plasticity and variation in adult body size (Weishampel, 1986; Sander and Klein, 2005; Klein and Sander, 2007; see Moser, 2003 and Klein and Sander, 2007 for the influence of taphonomic processes on these size estimates). Seebacher (2001) gave the highest mass estimate with a body mass at 1072 kg for an individual of 6.5 m length, and Sander (1992) estimated an 8 m individual at nearly 2200 kg. Recent CAD-model based estimates vary between 600 and 838 kg, depending on the overall density and amounts of soft tissues assumed, for an individual of roughly 6 m total length (Mallison, 2010a, 2011a).
The Locomotion Controversy
Different locomotion modes have been debated in the literature for Plateosaurus engelhardti. Variations of a lizard-like locomotion were assumed by Jaekel (1910) and Fraas (1913). Jaekel soon revised his opinion and proposed a kangaroo-style hopping (Jaekel, 1911, 1913), a change of heart for which Tornier (1912) viciously attacked him. Several authors concluded or implied that P. engelhardti was exclusively (Wellnhofer, 1994) or obligatorily quadrupedal (Paul, 1987, 1997, 2000), or was facultatively bipedal at high locomotion speeds (Galton, 1976, 1990; Weishampel and Westphal, 1986; Christian et al., 1996; van Heerden, 1997; Moser, 2003; Galton and Upchurch, 2004). Von Huene's (1926) conclusion that P. engelhardti was an obligate biped was reaffirmed by Bonnan and Senter (2007) and Rauhut et al. (2011). A detailed assessment of the locomotion of Plateosaurus conducted using computer-aided engineering (CAE) provided new approaches for determining body mass, motion range, posture and, therefore, locomotion mode (Mallison, 2010a, 2010b, 2011a). Quadrupedal reconstructions of Plateosaurus engelhardti in the form of skeletal mounts, models and drawings were mostly found to have incorrect limb proportions and disarticulated joints, usually the wrist and elbow (Mallison, 2010b). Correctly mounted, the forelimb is only about half as long as the hind limb, unsuitable for locomotion (Mallison, 2010a). Furthermore, the motion range of the forelimb under load is restricted, again indicating that the forelimb did not play a significant role in locomotion (Mallison, 2010b). Digital 3D assessment of the articulation of radius and ulna using similar techniques to those employed here confirmed the conclusions of Bonnan and Senter (2007) that pronation was possible to a very limited degree only, insufficient for locomotion (Mallison, 2010a, 2010b, 2011a). Further indications on the locomotion mode of Plateosaurus engelhardti can be expected from the anatomy of the manus. The pentadactyle hands have robust digits I through III with claw-like unguals and more delicate and slender digits IV and V, the latter not orientated parallel to the other digits, but angled ulnary (Mallison, 2010a, 2010b). This hand was interpreted as a grasping organ by von Huene (1926).
Study Objectives
Aims of work. It is the aim of this study to investigate the capabilities of the hands of Plateosaurus engelhardti with regards to locomotion by determining the motion ranges of the individual joints as well as of the manus as a whole in a CAE program based on NASTRAN (NASA Structural Analysis System). For this purpose, the ability to evenly hyper-extend the digits is of key importance, as is the ability of the metacarpus to support the weight of the animal. Below we formulate two hypotheses on these conditions. We also describe the flexion limits of the digits, but refrain from a detailed interpretation, as it would require a detailed comparison to other taxa what is beyond the scope of this paper.
Definition: ONP (osteologically neutral pose). The term ONP is used to describe the position in which the long axes of two bones articulating with each other are approximately parallel to each other in lateral view, as they are in a human hand placed flat on a table with the fingers straight. In dorsal view, ONP means a position in which the articulation surfaces in this laterally determined ONP position show the best fit (in humans this equals a position with relaxed, not laterally or medially flexed fingers).
Hypothesis 1: Plateosaurus engelhardti was able to hyper-extend its digits significantly. Quadrupedal locomotion with forelimbs much shorter than the hind limbs requires a significant amount of hyper-extendibility of the digits to reach an adequate stride length even for slow efficient locomotion. This hyper-extension ability is tested in case the conclusions of Bonnan and Senter (2007) and Mallison (2010a, 2010b, 2011a) were in error, and pronation of the lower arm was possible to a degree that allowed the palm to face the ground. Hyperextension of all digits that are involved in support must be even, as the motion limit of the whole hand is defined by the least mobile digit.
Hypothesis 2: The sum of the shaft cross section of radius and ulna of Plateosaurus engelhardti were approximately identical to that of all metacarpals potentially involved in locomotion. Given the extreme difference in length between the forelimb and hind limb in Plateosaurus it is plausible to assume that the hand had a digitigrade posture if it was used during locomotion. In that case, metacarpus and antebrachium carried equal amounts of body weight and were stressed across similar motion angles, and thus were subjected to similar forces. Therefore, the cross section areas of the bones should be nearly identical. A plantigrade (or semiplantigrade, with a large, weight-carrying heel) posture would lead to lower cross sections in the metacarpals, as part of the weight would be carried by the carpus, which would come into contact with the ground. Conversely, forces and thus cross section areas may differ significantly if force projection onto objects during other actions (e.g., grasping) created the main forces on the bones.
These hypotheses can be tested by a classic, non-digital motion range study and bone measurements. Using digital files in a CAD program makes these tasks easier, because extensive support for the bones is not required (see Mallison, 2010a, 2010b, 2011a). However, using the 3D digital models in a motion modeling program allows even easier judgment of the effect of motion in many joints at the same time, including combinations of intermediate positions, not just maximum flexion and extension. Also, results can be visualized with ease, by exporting still frames or videos of the main modeling window, which show motion sequences and positions in any desired view.
Abbreviations
PLSST – Paläontologische Lehr- und Schausammlung, Fachbereich Geowissenschaften, Eberhard Karls Universität Tübingen, Eberhard-Karls-University Tübingen (formerly IFGT or GPIT)
MfN – Museum für Naturkunde – Leibniz Institute for Research on Evolution and Biodiversity at the Humboldt University Berlin
ONP – osteologically neutral posture
SMNS – Staatliches Museum für Naturkunde Stuttgart
YPM – Yale Peabody Museum.
MATERIAL
Fossil Material
The left manus of Plateosaurus engelhardti "GPIT Skelett 2" was CT scanned by B. Ludescher (University Hospital, Eberhard-Karls-University Tübingen), with a slice thickness of 1 mm and slice overlap of 0.5 mm. File extraction was performed in AMIRA 4.0 by HM (see Mallison, 2010a) (note: the specimen is not officially numbered, but "GPIT Skelett 2" or "GPIT skeleton 2" was previously used in the literature).
 "GPIT Skelett 2" is mounted in the PLSST (Figure 1). The mount consists of two individuals, which were founded in Trossingen, Germany, during the PLSST's 1921-1923 excavations. Details on which parts belong to which individual can be found in von Huene (1932). Of the left hand, assigned by von Huene to Plateosaurus quenstedti (also a junior synonym of P. engelhardti, see Galton, 1984; Moser, 2003), are preserved, distal to the wrist: metacarpals I through V, complete digits I, II and III, and incomplete digits IV and V (Figure 1). The distal phalanges of digit IV are badly preserved and were not separated during preparation, and were extracted from the CT scans as a single object. Therefore, only the two proximal joints of digit IV could be used for the simulation. However, this still allows an acceptable approximation of the mobility of the digit for the purpose of this study, due to the shortness of the elements and the large uncertainties of motion range in all its joints. In digit V only the basal phalanx is preserved. Because of the assumed functional importance of digit V it was necessary to derive a better approximation of the actual conditions than possible by omission of the missing elements. Therefore, a scaled-down digital copy of the basal phalanx was used as substitution for the second phalanx. The presence and size of the missing phalanges is known from other finds, including MFN skeleton C. For the joint between mPh-V-1 and mPh-V-2, the preserved distal articular surface of mPh-V-1 offers valid information about joint morphology. Information on the more distal joints is lacking. Therefore, we desisted from substituting the other missing distal phalanges as this would have introduced highly speculative data. Also, not substituting the missing bones with copies of those of other individuals guaranteed that all bones come from the same individual, and therefore definitely from the same species and the same ontogenetic state. By keeping both digits IV and V intentionally reduced compared to better preserved specimens our model can account for potential intraspecific variability, which could be the cause for the much reduced preservation in the GPIT skeleton 2 specimen. This means that results here are valid as minimum scenarios. For hyper-extension the reduction has no bearing, because digits IV and V are significantly weaker than digits I through III, and thus likely played only a minor role in locomotion, if at all.
"GPIT Skelett 2" is mounted in the PLSST (Figure 1). The mount consists of two individuals, which were founded in Trossingen, Germany, during the PLSST's 1921-1923 excavations. Details on which parts belong to which individual can be found in von Huene (1932). Of the left hand, assigned by von Huene to Plateosaurus quenstedti (also a junior synonym of P. engelhardti, see Galton, 1984; Moser, 2003), are preserved, distal to the wrist: metacarpals I through V, complete digits I, II and III, and incomplete digits IV and V (Figure 1). The distal phalanges of digit IV are badly preserved and were not separated during preparation, and were extracted from the CT scans as a single object. Therefore, only the two proximal joints of digit IV could be used for the simulation. However, this still allows an acceptable approximation of the mobility of the digit for the purpose of this study, due to the shortness of the elements and the large uncertainties of motion range in all its joints. In digit V only the basal phalanx is preserved. Because of the assumed functional importance of digit V it was necessary to derive a better approximation of the actual conditions than possible by omission of the missing elements. Therefore, a scaled-down digital copy of the basal phalanx was used as substitution for the second phalanx. The presence and size of the missing phalanges is known from other finds, including MFN skeleton C. For the joint between mPh-V-1 and mPh-V-2, the preserved distal articular surface of mPh-V-1 offers valid information about joint morphology. Information on the more distal joints is lacking. Therefore, we desisted from substituting the other missing distal phalanges as this would have introduced highly speculative data. Also, not substituting the missing bones with copies of those of other individuals guaranteed that all bones come from the same individual, and therefore definitely from the same species and the same ontogenetic state. By keeping both digits IV and V intentionally reduced compared to better preserved specimens our model can account for potential intraspecific variability, which could be the cause for the much reduced preservation in the GPIT skeleton 2 specimen. This means that results here are valid as minimum scenarios. For hyper-extension the reduction has no bearing, because digits IV and V are significantly weaker than digits I through III, and thus likely played only a minor role in locomotion, if at all.
Computer Programs
Computer Aided Design. 'Rhinoceros 4.0 NURBS modeling for Windows®' (henceforth 'Rhino'; www.rhino3d.com/) by Robert McNeel & Associates (www.en.na.mcneel.com/) is a CAD program. Rhino was used to test and improve the digital articulation of the bones files.
Kinematic modeling. NX5 by Siemens AG is a Computer-aided Engineering (CAE) program capable of fully kinetic-dynamic modeling. The program is limited to modeling rigid bodies, so that flexible bodies can only be simulated by connecting series of rigid bodies. However, for this study deforming bodies were not required. With respect to accuracy and speed, NASTRAN-based programs such as NX5 can be regarded as intermediate between simple biomechanical calculation and highly detailed musculoskeletal modeling. However, some aspects such as collision conditions are difficult to define appropriately (Mallison, 2011a). For further details on NASTRAN-based modeling see Mallison (2010a, 2011a, 2011b).
METHODS
Digital Skeleton Articulation
The digitized files of the bones were articulated in Rhino in order to reconstruct the manus, using the methods employed before by Mallison (2010a, 2010b, 2010c). The configuration was based on articulated fossil finds, e.g., SMNS F33, and 'best fit' of the bones. A roughly articulated version of the manus (see Mallison, 2010a, 2010b) was used as the basis for this study. The articulation was checked for errors, which were corrected by altering the 3D association of the individual bone files (see below). This was achieved by rotating and sliding bones individually and in groups and reviewing the resulting arrangement in six axial views and a freely rotating view, all in parallel projection mode. 'Best fit' was determined on the basis of the preserved proximal and distal facets of the bones. The lack of knowledge on the exact shapes and thicknesses of soft tissues involved in forming the joints leads to uncertainties in the results, which are discussed below.
For transfer of the data into the kinetic/dynamic modeling program, each bone was exported as a separate polygon mesh (STL) file.
Motion Range Modeling
The placement of bone files in Rhino defines their location in NX5, so that no further corrections on position had to be undertaken. After import of the raw bone data, the modeling in NX5 consisted of three steps:
1. Definition of suitable settings for the simulation.
2. Modeling the digital joints.
3. Modeling grasping scenarios with generic bodies
4. Documentation of the simulation data.
1. Definition of suitable settings for the simulation. The general settings for a simulation should always be as simple as possible, avoiding the introduction of errors and making simulation results easier to compare to other studies. Gravity, for example, is not relevant in this study and was therefore switched off. Euler integration with a frame rate of 50 s-1 with two integration steps per frame guaranteed a sufficiently accurate display. Although it is far less accurate than other options available in NX5 using integration with variable time steps, for the purpose of this study Euler integration is sufficient, and reduces calculation time significantly.
The study did not include a stress analysis, so it was possible to give the bones unrealistically low density values. This further reduced calculation times for simulations, because the forces caused by inertia are minimal.
2. Modeling the digital joints. The basic motions, maximum flexion and maximum hyper-extension of the digits, were modeled via sinusoid functions. To simplify the modeling process and save computation time, flexing and extending motions were modeled in separate simulation files.
Using motion modeling instead of simple static analysis of maximum flexion and extension positions shows the posture of the entire digit or manus throughout motion in all joints. Joints with complicated geometries (i.e., those that differ significantly from a single-axis rotary joint) could be judged and adjusted much easier when shown in a fluid motion than in static poses. Also, motion allowed better judgment if the determined motion ranges for the individual joints added up to a consistent whole-digit posture.
3. Documentation of the simulation data. The results of the simulation were documented as follows:
a) The motion of the distal bone in relation to the proximal bone of each joint was plotted in-program as graphs.
b) Additionally tables were exported giving joint axis orientation and displacement per time.
CAVEATS
Remarks on the Metacarpus
The metacarpals were regarded as an immobile block in the simulations. There are a number of indications that the metacarpals could move little or not at all against each other. The presence of planar lateral contact surfaces is inconsistent with motion. The configuration of the metacarpals in articulated fossils supports this interpretation. Specimens SMNS 13200k, SMNS F 33 and MSF 23 preserve articulated hands with a compact, parallel metacarpal placement, which is also seen in other "prosauropod" genera such as Seitaad ruessi Sertich and Loewen, 2010 and Sarahsaurus aurifontanilis Rowe et al., 2010.
Articular Cartilage
A factor significantly influencing the motion range of a joint is the thickness and shape of the articular cartilage. Several authors have provided evidence that dinosaurs or archosaurs developed thick cartilage caps at the articular surfaces (Schwarz et al., 2007; Bonnan et al., 2013; Holliday et al., 2001, 2010; Mallison, 2010b, 2010c). The bone configuration of the manus of Plateosaurus engelhardti from Mallison (2010b) showed minor contact or overlap between various neighboring bones. In some cases, moving the bones distally to eliminate overlap caused significant gaps to open between articulation surfaces. These gaps may indicate thick articular cartilage in those cases where they are not caused by postmortal damages.
Usually, the corresponding functional articular surfaces fit each other in living animal without large gaps, whereas this is not the case in Plateosaurus. Also, in mammals, the bony condylar morphology reflects the morphology of the functional articular end, unlike in alligators and other archosaurs (Haines 1938, 1969; Holliday et al. 2010). In the manus of Plateosaurus engelhardti, there are several joints in which significant differences exist, e.g., a deep groove between the condyles of the distal articular surfaces is not countered by a prominent keel between the facets of the proximal articular surfaces. This is the case, e.g., in the joints between the ungual phalanges of digits I through III and their next proximal phalanges. Smaller differences between corresponding articular surfaces are also found in the joints MC I/mPh-I-1, MC II/mPh-II-1 and mPh-III-1/mPh-III-2. The exact thickness of the cartilage in each separate digit joint cannot be determined. In situ finds, however, indicate that cartilage thickness was not significantly larger than in extant reptiles. Additionally, Bonnan et al. (2013) found that "thinner articular cartilage would appear to be associated with a subchondral bone shape displaying well-developed surfaces, condyles and convexity. In contrast, [...] in archosaurs, thicker articular cartilage is associated with flatter, poorly-developed surfaces and a less convex shape." Where possible, we therefore assumed thin, mammal- and extant reptile-like cartilage caps if the articular ends of a bone were well-ossified with distinct, well-developed articular facets. This is in contrast to the ends of longbones, which in dinosaurs were covered by thick cartilage layers (see Holliday et al., 2010, Mallison, 2010a, 2010b, 2011a; Bonnan et al., 2013). Where the articular ends were smoothly rounded as in the phalanges of digits IV and V, the amount of cartilage must remain speculative.
This approach guarantees that we do not underestimate range of motion by virtually "pressing" bone on bone, but do not induce unnecessary speculation.
Postmortem Damages
Some bones of the hand of GPIT skeleton 2 show taphonomic deformation. The proximal articular surface of mPh-I-2 is slightly less broad than the distal articular surface of mPh-I-1, and the distal end of the phalanx looks pinched in dorsal view. This indicates a possible deformation of the ungual. In other specimens the ungual indeed is slightly broader, but this difference may also have been caused by variation.
The distal articular surface of mPh-II-1 is remarkably wide compared to the proximal articular surface of mPh-II-2, indicating a mediolateral crushing of the latter. Again, other specimens show a differing morphology, with the articular faces roughly equally wide.
A break-like structure in the middle of mPh-III-3 and the slightly oblique medial condyle of the distal articular surface of mPh-III-3 indicate taphonomic damage. MPh-IV-2 is poorly preserved and probably damaged. However, these deformations are not so large as to make a cautious simulation of the motions impossible, as the results are probably only slightly influenced. Comparison to other specimens indicates that our results are correct.
RESULTS
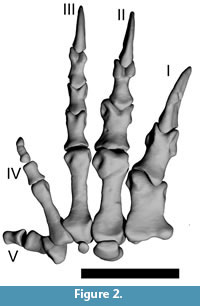 Articulation
Articulation
Compared to the initial version of the articulated manus from Mallison (2010a), minor adjustments were required to correct for impossible or unlikely articulations, e.g., if bones overlapped. MPh-II-2 was upside down in the initial version (Mallison, personal commun., 05/2010, however this was corrected before publication of Mallison [2010a]), and was rotated 180° to correct. Figure 2 shows the digitally articulated hand in neutral pose (straight digits) in dorsal view.
The asymmetric development of the manus suggests partitioning of the digits into two groups, which are easily recognizable by differences in overall morphology: digits I through III and digits IV and V. This morphological grouping is confirmed by the motion range analysis.
Digits I – III. (Figure 3, Figure 4, Figure 5, Figure 6, Figure 7) In neutral position (neither flexed nor hyper-extended), the long axis of digit I is angled 10° medially compared to the midline of MC III. Digit II and III are straight and orientated nearly parallel, which is also the case in all other examined specimens. In this context it is worth noting that in the feet of individuals assigned to Plateosaurus, there are marked differences, with some individuals having straight toes, whereas the pedal digits of others are markedly curved laterally.
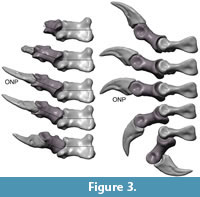 The distal articular surface of MC I through III differ somewhat in shape. The medial condyle of MC I is much smaller than the lateral condyle, whereas in MC II it is only slightly smaller. The median sulcus between the condyles of MC I is deeper than that of MC II. The distal articular surface of MC III has a slightly convex shape without recognizable condyles or median sulcus. It is broad, extends slightly laterally and curves slightly proximally. All metacarpals have large collateral ligament pits.
The distal articular surface of MC I through III differ somewhat in shape. The medial condyle of MC I is much smaller than the lateral condyle, whereas in MC II it is only slightly smaller. The median sulcus between the condyles of MC I is deeper than that of MC II. The distal articular surface of MC III has a slightly convex shape without recognizable condyles or median sulcus. It is broad, extends slightly laterally and curves slightly proximally. All metacarpals have large collateral ligament pits.
The preungual phalanges of digits I – III are uniform in morphology and generally show the following features: The proximal articular surface has moderately to well-developed lateral and medial cotyles with a median keel. The distal articular surface has well-developed lateral and medial condyles and a distinct median sulcus, so that the overall appearance is spool-shaped. There are distinct collateral ligament pits, with varying degrees of excavation. The proximal ends of the phalanges show dorsal and ventral processus for the attachment of the flexor and extensor tendons. The ventral processus is usually larger than the dorsal processus. More details on the features of the individual phalanges will be given below. The unguals of digits I – III have proximal articulation surfaces that greatly resemble those of the more basal phalanges. Overall, the unguals are slightly curved, laterally compressed and show a distinct lateral groove.
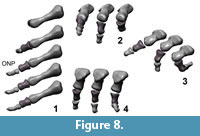
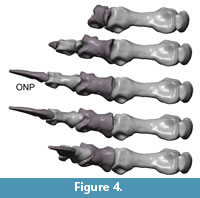 Digits IV – V. (Figure 8) In digits IV and V the articular surfaces have no prominent keels, sulci, condyles or cotyles. The joints are simple spherical joints, limited only by prominent rims around the facet, or have plain articular surfaces. The long axis of digit III points 35° ulnary compared to the midline of MC III. The long axis of digit V points nearly 90° ulnary and 30° palmary compared to the midline of MC III. Collateral ligament pits are small and shallow or not visible. Proximal dorsal and ventral processus are not developed.
Digits IV – V. (Figure 8) In digits IV and V the articular surfaces have no prominent keels, sulci, condyles or cotyles. The joints are simple spherical joints, limited only by prominent rims around the facet, or have plain articular surfaces. The long axis of digit III points 35° ulnary compared to the midline of MC III. The long axis of digit V points nearly 90° ulnary and 30° palmary compared to the midline of MC III. Collateral ligament pits are small and shallow or not visible. Proximal dorsal and ventral processus are not developed.
Bone Diameters
The minimal shaft cross sections of metacarpals I through III are roughly 90% (8.73 cm2) of those of radius and ulna (9.48 cm2). The ratio increases to nearly 1:1 if cross sections are measured at the articular ends (23.56 cm2 vs. 24.12 cm2).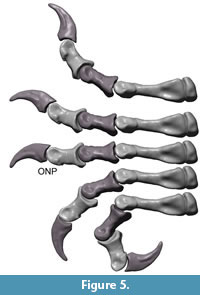 In contrast, if only metacarpals II and III are considered the ratio decreases to just below 50% for the minimal shaft diameters (4.43 cm2 vs 9.48 cm2), and ~55% for the diameters of the articular ends (13.66 cm2 vs. 24.12 cm2).
In contrast, if only metacarpals II and III are considered the ratio decreases to just below 50% for the minimal shaft diameters (4.43 cm2 vs 9.48 cm2), and ~55% for the diameters of the articular ends (13.66 cm2 vs. 24.12 cm2).
Digit Mobility
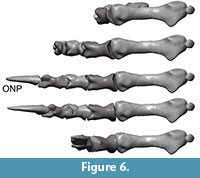
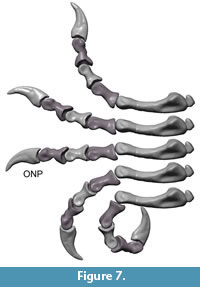
Flexion and hyper-extension angles in relation to the long axis of the manus for all phalanges are given in Table 1. Figure 3, Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8 show views of the individual digits flexing and extending.
Digits I – III. The well-developed prominent condyles, proximal dorsal and ventral processus restrict the flexibility of the digits. The flexion of each joint reaches higher degrees than the hyper-extension.
 In maximum flexion, the ungual of digit I reaches an angle of 151° in relation to the long axis of the manus, while the lateral divergence of digit I from the midline of manus (long axis of MC III) is reduced to less than 2°. In consequence of this orientation digit I cannot be opposed. The ungual of digit I does not reach the palm. The ungual reaches a higher degree of flexion at nearly 80° than the basal phalanx.
In maximum flexion, the ungual of digit I reaches an angle of 151° in relation to the long axis of the manus, while the lateral divergence of digit I from the midline of manus (long axis of MC III) is reduced to less than 2°. In consequence of this orientation digit I cannot be opposed. The ungual of digit I does not reach the palm. The ungual reaches a higher degree of flexion at nearly 80° than the basal phalanx.
At maximum hyper-extension, an angle of nearly 50° in relation to the long axis of the manus is reached by the ungual of digit I. Digit I rotates to a slightly medial orientation due to rotation of mPh-I-1 around its long axis, and the medially tilt of digit I reaches 29°.
Digit II and digit III remain nearly parallel in flexion and hyperextension. Maximal flexion angles the ungual of digit II against the metacarpus by 189°, but the ungual does not or only barely reach the palm. The highest flexion degree of a distal bone against a proximal bone in digit II is 80° (ungula mPh-II-3 against mPh-II-2). Digit II is not very flexible in extension. In relation to the long axis of the manus the ungual is angled 84° at maximum hyperextension of the digit.
 In maximum flexion, the ungual of digit III is angled against the metacarpus by 237° and the tip of the ungual reaches the palm. Maximum flexion of the distal phalanx in relation to the proximal bone reaches between 50° (the two proximal joints) and 70° (the two distal joints) in digit III. MPh-III-1 can perform ulnar rotation during flexion. This rotation causes an increase of medial deflection of the ungual to nearly 7° in relation to the long axis of the manus at maximum flexion. At this angle, its medial orientation places the ungual of digit III nearly into contact with that digit II when both digits are strongly flexed.
In maximum flexion, the ungual of digit III is angled against the metacarpus by 237° and the tip of the ungual reaches the palm. Maximum flexion of the distal phalanx in relation to the proximal bone reaches between 50° (the two proximal joints) and 70° (the two distal joints) in digit III. MPh-III-1 can perform ulnar rotation during flexion. This rotation causes an increase of medial deflection of the ungual to nearly 7° in relation to the long axis of the manus at maximum flexion. At this angle, its medial orientation places the ungual of digit III nearly into contact with that digit II when both digits are strongly flexed.
In hyperextension, digit III is the most flexible of the first three digits. The basis joint reaches 40° when fully extended, so that the tip of the ungual is angled against the metacarpus by 104°.
Digits IV – V. The maximum flexion of the basal joint of digit IV reaches 80° and the flexion of joint mPh-IV-1/mPh-IV-2 reaches 60°. Flexed, the digit is orientated medially and may have contacted the palm. Digit IV is less flexible in hyperextension. It reaches 15° in the basis joint and 20° in the joint between mPh-IV-1 and mPh-IV-2.
Similar to digit III, digit IV can rotate slightly ulnary by nearly 7° during flexion and in neutral position. This rotation causes a more medial deflection of the distal phalanges. Additionally, mPh-IV-1 can rotate ulnary around the sagittal axis by 20° when hyper-extended.
Digit V is the most flexible digit. In maximum flexion both modeled joints reach 90° and the digit opposes digit III, but was probably not able to oppose to digits I and II because of its short length. Digit V seems to have been able to reach the palm. In hyperextension, the digit is more flexible than digit IV. The joint between mPh-V-1 and mPh-V-2 reaches the largest hyper-extension angle of the entire hand (50°). However, the high values for digits IV and V must be regarded with caution, because the lack of well-defined articular surfaces forces speculation into the assessment of mobility. Extension of the unguals of digit IV and V cannot be assessed for GPIT skeleton 2 due to their incomplete preservation.
DISCUSSION
 Testing the Hypotheses
Testing the Hypotheses
Hypothesis 1: Plateosaurus engelhardti was able to hyper-extend its manual digits significantly. In most joints, large hyper-extension angles are prevented by the morphology of the articulation surfaces. A significant and uniform hyper-extension of all digits is prevented by the differing values for maximum extension, indicating that the manus did not allow for large stride lengths. Not even the two longest digits II and III show a uniform pattern of extension.
- Digit I has limited mobility in the basal joint during extension and cannot reach significant hyper-extension values. The slightly medial orientation of the digit during extension and its shortness also indicate that the first digit was not able to support body weight in a quadrupedal posture, as it would not touch the ground at a significant length.
- Digit II has a similarly limited ability to hyper-extend in the basal joint as digit I. Due to a higher number of distal segments and a significantly more flexible joint between mPh-II-1 and mPh-II-2 than in the corresponding joint of digit I, digit II as a whole is more flexible than digit I during hyper-extension. Although digit II is also longer than digit I, it is neither as long nor as flexible as digit III, and thus was not capable of forming a congruent support during locomotion together with digit III.
- Digit III is more mobile in the basal joint than digit I and II, and was long enough to be used effectively in quadrupedal locomotion. However, it is not significantly stronger than digit II in cross section area, and would alone not have been strong enough to support a significant part of the body weight. Additionally, the distal joints of digit III are limited in their mobility during hyper-extension.
- Digit IV is the least extensible digit. It has greater lateral flexibility than the other digits, but is incapable of forming a useful support due to its lateral orientation.
- Digit V has a greater flexibility during hyper-extension than digit IV, but it is quite unsuited for quadrupedal locomotion due to its lateral orientation, which prevents ground contact.
In summary, the mainly small hyper-extension angles possible do not match an adaptation to greater stride length in the forelimb, which seems necessary for quadrupedal locomotion. The small hyper-extension angles of the basal joints (except in digit III) also hinder a plantigrade gait. A digitigrade use of the manus during active locomotion is not likely due to the low dorsal mobility within the digits. Only stabilizing of the anterior body during feeding or drinking at ground level seems possible, with laterally directed digits and medially facing palms. Consequently, Hypothesis 1 is not confirmed.
Hypothesis 2: The sum of the shaft cross section of radius and ulna of Plateosaurus engelhardti were approximately identical to that of metacarpals I through III. The only digits long and mobile enough to bear significant weight in a digitigrade posture are digits II and III. The cross section areas of their metacarpals are roughly half those of radius and ulna, indication that the manus was not used to support the body during locomotion.
In contrast, the cross sections of all metacarpals together closely match those of radius and ulna. Therefore, it seems likely that significant forces acted in the hand as a whole, during other actions than locomotion.
Comparison to Other Extinct Taxa
Basal sauropodomoprhs ('prosauropods'). Many basal sauropodomorphs are incompletely known, and those lacking a manus (e.g., Anchisaurus including Ammosaurus, Ruehleia, Pantydraco [Thecodontosaurus] caducus (Yates, 2003b), Chromogisaurus (Ezcurra, 2010) and Saturnalia (Langer et al., 2007)) or phalanges in the variably developed digits IV and V, as is the case in Thecodontosaurus antiquus YPM 2195 (Benton et al., 2000), are obviously unsuitable subjects for comparison here.In the following, we only mention a selection of taxa we studied ourselves, or for which sufficient data on the manus was either published.
In the basal sauropodomorph Seitaad the distal end of MC I is broader and sturdier than in Plateosaurus. The torsion of digit I is similar in the two taxa. Sertich and Loewen (2010) note that in Seitaad "digit IV preserves three complete phalanges. The first two are similar in overall morphology to the proximal phalanges of digits II and III, though with shallow collateral pits and deeply divided distal condyles." The well-developed two basal phalanges contrast with the strong reduction seen in Plateosaurus. Similarly, "the distal phalanx of digit V is much longer than the proximodistally short distal phalanges of other basal sauropodomorphs including Efraasia, Massospondylus, and Plateosaurus" (Sertich and Loewen, 2010).
The metacarpals of Seitaad, based on Sertich and Loewen (2010, figure 7) and own photographs, seem to form an arc reminiscent of that in sauropods, in contrast to the nearly flat metacarpus of Plateosaurus. Measuring the length ratio of metacarpals IV to III on Sertich and Loewen (2010, figure 7), an admittedly imprecise measurement, indicated that the ratio is roughly 0.84, whereas in Plateosaurus it is around 0.74. Overall, the metacarpus and digit development of Seitaad therefore resembles a sauropod manus, and its adaptation to weight bears much more than the manus of Plateosaurus does. This resemblance adds slight biomechanical support to the phylogenetic placement of Seitaad much closer to sauropods than Plateosaurus (Sertich and Loewen, 2010, figure 12A) based on an improved version of the matrix of Yates (2007) rather than a placement close to Plateosaurus, as Sertich and Loewen recover using the matrix of Upchurch et al. (2007).
Massospondylus carinatus has a metacarpal IV to III ratio of ~0.82 (data from Cooper, 1981), intermediate between Seitaad and Plateosaurus. Otherwise its hand is overall similar to that of both taxa (based on figures in Cooper, 1981). Like the manus of Efraasia (figure 9 in Galton, 1973), the manus of Adeopapposaurus apparently is highly similar in overall digit proportions and phalangeal proportions to that of Plateosaurus (based on Martínez, 2009, figures 18, 19). Martínez (2009) recovered the two as close relatives. Sertich and Loewen's (2010) analyses, however, do not show an especially close relationship between the two taxa, independent of the matrix used. The instability of cladistic analyses of the "prosauropods" may indicate that a large number of character states independently evolved convergently; if so, adaptations of the manus for locomotion or, conversely, other functions such as grasping would be likely candidates. Our results, and even a comprehensive study of all available "prosauropod" manus, are unlikely to shed light on interrelationships of basal sauropodomorphs. Efraasia, however, is basal to the difficult to resolve group of "prosauropods" in Sertich and Loewen (2010), independent of the matrix they use, which can be seen as an indication that a Plateosaurus-like manus was an ancestral characteristic that was subsequently lost once or repeatedly.
The manus of more derived (according to Sertich and Loewen (2010)) taxa like Melanorosaurus is difficult, due to the lack of well-preserved remains. Of Antetonitrus only MC I and II are known, but these two bones are already much broader (sturdier) than in Plateosaurus (Yates and Kitching, 2003), and therefore adapted to supporting the animal during locomotion. The manus of Melanorosaurus shows reduction of the phalanges of digits II and III akin to later sauropods, a clear adaptation to weight bearing.
Ornithischia. Here, only some facultatively or obligatorily bipedal ornithischians will be discussed. All ornithischians are considered to have been herbivorous and in this respect similar to Plateosaurus engelhardti. Also similar to Plateosaurus, most bipedal ornithischians have relatively broad hands, mostly with all five digits developed. Some ornithopods have apparently specialized hands, partly with remarkable superficial similarities to Plateosaurus.
The hand of the basal ornithischian Heterodontosaurus is overall the most similar among non-"prosauropod" dinosaurs to that of Plateosaurus. The asymmetric development of the digits, the shapes of the articular surfaces and the robust claws on the first three digits (Santa-Luca, 1980; Norman et al., 2004) are reminiscent of the respective features of Plateosaurus. This may indicate a similar use of the hand as a grasping organ in Plateosaurus. However, the phalanges of digitis I-III of Heterodontosaurus are proportionally significantly longer and more slender than those of Plateosaurus, and the difference in length between the two groups of digits thus decidedly more pronounced. The more robust development of the manus of Plateosaurus may indicate adaptation to the production of larger forces. Overall, the similarities appear to support a non-locomotory adaptation of the manus in Plateosaurus. In contrast, the manus of Tenontosaurus Ostrom, 1970, despite showing a slight partitioning of digits into two groups as well, is significantly more massive, the metacarpals are arranged in an arc, and digits IV and V are better ossified than in Plateosaurus. All these are adaptations for weight bearing, matching the proportionally longer forelimb.
Similarly, the hyper-phalangy of digit V in Iguanodontia (Norman, 2004) and the ability to greatly hyper-extend the digits and other specializations for quadrupedal locomotion in digits II through IV in Iguanodontia (Norman, 2004; Carpenter and Wilson, 2008) are clear discrepancies to Plateosaurus. These differences are concordant with the hypothesis that Plateosaurus was obligatorily bipedal.
Summary. The comparison to other (facultatively) bipedal dinosaur taxa reveals that in some respects the hand of Plateosaurus is more reminiscent to the hands of bipedal ornithischians than those of the more closely related sauropodomorphs. It can therefore be reasoned that Plateosaurus engelhardti had a manus which was in its basic design characteristic for a large bipedal herbivore – broad and potentially with prehensility. Such a manus may have been typical of basal sauropodomorphs.
Ontogenetic Development of Bipedality
Our conclusion that the hand of Plateosaurus was unsuited to play any role in locomotion fits those of Bonnan and Senter (2007) and Mallison (2010a, 2010b) that posture and limb proportions of Plateosaurus made quadrupedal locomotion impossible in large sub-adults and adults. Hatchlings, however, probably had a different locomotion mode, using a crawling quadrupedal gait. The hatchlings of the proportionally similar and closely related sauropodomorph Massospondylus were quadrupedal (Reisz et al., 2005), and it is likely that Plateosaurus and other similar basal sauropodomorphs also had proportionally large heads and an anterior center of mass position as hatchlings. Because the adults were bipedal, an ontogenetic change of the locomotion mode must have occurred (Reisz et al., 2005; Bonnan and Senter, 2007). Possibly, the hatchlings moved in a manner similar to that of lizards, with a subhorizontal orientation of the humerus and sideways orientated hands (Bonnan and Senter, 2007). Alternatively, the hatchlings may have been capable of limited pronation of the manus, and this capability was lost during growth (Bonnan and Senter, 2007). How the ontogenetic transition occurred alongside the changes in body, limb and skull proportions is unclear, because of the lack of fossils of young individuals of Plateosaurus (Weishampel and Westphal, 1986; Sander, 1992). A gradual development of, e.g., a pronounced grasping capability during ontogeny is plausible, with the change from quadrupedal to bipedal locomotion freeing the hand from supporting the body. On the other hand, digits II and III may well have been strong and mobile enough to support the body weight of a hatchling moving slowly, so that there is no reason to assume an ontogenetic shift in digit proportions.
CONCLUSIONS
The robust digits I through III, the stout metacarpus and the very robust bones of the forelimb indicate that Plateosaurus was able to exert large forces with the manus. There is a distinct split into three function digit groups, with digit I short and powerful, digits II and III long and powerful, and digits IV and V much reduced. However, the manus was not adapted to locomotion, because those digits long and strong enough to play a role in supporting the body (II and III) were together not strong enough to support as much weight as either radius and ulna or the metacarpus, and did not hyper-extend uniformly and sufficiently. As a sub-adult and adult, Plateosaurus engelhardti was obligatorily bipedal. How the shift from likely quadrupedal hatchlings to bipedal adults occurred is unclear, as there are no fossils known of young of Plateosaurus. The manus of Plateosaurus may have been well-adapted to grasping, but further work is required.
ACKNOWLEDGEMENTS
N. Klein (Steinmann-Institut, Bonn, Germany) and D. Schwarz-Wings (MfN Berlin, Germany) supported the author with feedback and access to literature. J, Sertich (Stony Brook University, Stony Brook, New York, USA) provided pictures of Seitaad. S. Schliwa (Anatomisches Institut, Bonn, Germany) granted access to a manus of Homo sapiens. Kind reviews of the first draft by M. Belvedere (MfN Berlin, Germany) and M. Bonnan (Richard Stockton College, New Jersey) greatly improved this paper.
REFERENCES
Benton, M.J., Juul, L., Storrs, G.W., and Galton, P.M. 2000. Anatomy and systematics of the prosauropod dinosaur Thecodontosaurus antiquus from the Upper Triassic of Southwest England. Journal of Vertebrate Paleontology, 20:77-108.
Bonnan, M.F. and Senter, P. 2007. Were the basal sauropodomorph dinosaurs Plateosaurus and Massospondylus habitual quadrupeds?, p. 139-155. In Barret, P.M. and Batten, D.J. (eds.), Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (Special Papers in Palaeontology 77). Blackwell Scientific Publications, Oxford.
Bonnan, M.F., Wilhite, D.R., Masters, S.L., Yates, A.M., Gardner, C.K., and Aguiar, A. 2013. What lies beneath: sub-articular long bone shape scaling in eutherian mammals and saurischian dinosaurs suggests different locomotor adaptations for gigantism. PLOS ONE, 8(10):e75216. doi:10.1371/journal.pone.0075216.
Carpenter, K. and Wilson, Y. 2008. A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a biomechanical analysis of its forelimb. Annals of the Carnegie Museum, 76:227-263.
Christian, A., Koberg, D., and Preuschoft, H. 1996. Shape of the pelvis and posture of the hindlimbs in Plateosaurus. Paläontologische Zeitschrift, 70:591-601.
Cooper, M.R. 1981. The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance. Occasional Papers, National Museums and Monuments of Rhodesia, Series B, Natural Sciences, 6:689-840.
Ezcurra, M.D. 2010. A new early dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Argentina: a reassessment of dinosaur origin and phylogeny. Journal of Systematic Palaeontology, 8:371-425.
Fraas, E. 1913. Die neuesten Dinosaurierfunde in der schwäbischen Trias. Naturwissenschaften, 45:1097-1100.
Galton, P.M. 1973. On the anatomy and relationships of Efraasia diagnostica (HUENE) n. gen., a prosauropod dinosaur (Reptilia: Saurischia) from the Upper Triassic of Germany. Paläontologische Zeitschrift, 47:229-255.
Galton, P.M. 1976. Prosauropod dinosaurs (Reptilia: Saurischia) of North America. Postilla, 169:2-98.
Galton, P.M. 1984. Cranial anatomy of the prosauropod dinosaur Plateosaurus from the Knollenmergel (Middle Keuper, Upper Triassic) of Germany. I. Two complete skulls from Trossingen/Württ. With comments on the diet. Geologica et Palaeontologica, 18:139-171.
Galton, P.M. 1990. Basal Sauropodomorpha - Prosauropoda, p. 320-344. In Weishampel, D.B., Dodson, P., and Osmolska, H. (eds.), The Dinosauria. University of California Press, Berkeley.
Galton, P.M. 2001. The prosauropod dinosaur Plateosaurus Meyer, 1837 (Saurischia: Sauropodomorpha; Upper Triassic). II. Notes on the referred species. Revue de Paléobiologie, 20:435-502.
Galton, P.M. and Upchurch, P. 2004. Prosauropoda, p. 232-258. In Weishampel, D.B., Dodson, P., and Osmolska, H. (eds.), The Dinosauria, 2nd Edition. University of California Press, Berkeley.
Haines, R.W. 1938. The primitive form of epiphyses in the long bones of tetrapods. Journal of Anatomy, 72:323-343.
Haines, R.W. 1969. Epiphyses and Sesamoids, p. 81-115. In Gans, C. (ed.) Biology of the Reptilia. Academic Press, New York.
Holliday, C.M., Ridgely, R.C., Sedlmayer, J.C., and Witmer, L.M. 2001. The articular cartilage of extant archosaur limb bones: implications for dinosaur functional morphology and allometry. Journal of Vertebrate Paleontology, 21(Supplement to No. 3):62A.
Holliday, C.M., Ridgely, R.C., Sedlmayer, J.C., and Witmer, L.M. 2010. Cartilaginous epiphyses in extant archosaurs and their implications for reconstructing limb function in dinosaurs. PLOS ONE, 5(9):e13120. doi:10.1371/journal.pone.0013120
Jaekel, O. 1910. Die Fussstellung und Lebensweise der grossen Dinosaurier. Zeitschrift der Deutschen Geologischen Gesellschaft, Monatsberichte, 62:270-277.
Jaekel, O. 1911. Die Wirbeltiere. Eine Übersicht über die fossilen und lebenden Formen. Bornträger, Berlin.
Jaekel, O. 1913. Über die Wirbeltierfunde in der oberen Trias von Halberstadt. Paläontologische Zeitschrift, 1:155-215.
Klein, N. 2004. Bone histology and growth of the prosauropod dinosaur Plateosaurus engelhardti Meyer, 1837 from the Norian bonebeds of Trossingen (Germany) and Frick (Switzerland). Doctoral Thesis, Rheinische-Friedrich-Wilhelms-Universität Bonn, Bonn. http://hss.ulb.uni-bonn.de/2004/0466/0466.htm
Klein, N. and Sander, P.M. 2007. Bone histology and growth of the prosauropod dinosaur Plateosaurus engelhardti von Meyer, 1837 from the Norian bonebeds of Trossingen (Germany) and Frick (Switzerland), p. 169-206. In Barret, P.M. and Batten, D.J. (eds.), Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (Special Papers in Palaeontology 77). Blackwell Scientific Publications, Oxford.
Langer, M.C., Franca, M.A.G., and Gabriel, S. 2007. The pectoral girdle and forelimb anatomy of the stem-sauropodomorph Saturnalia tupiniquim (Upper Triassic, Brazil), p. 113-137. In Barrett, P.M. and Batten, D.J. (eds.), Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (Special Papers in Palaeontology 77). Blackwell Scientific Publications, Oxford.
Mallison, H. 2010a. The digital Plateosaurus I: body mass, mass distribution, and posture assessed using CAD and CAE on a digitally mounted complete skeleton. Palaeontologia Electronica, 13.2.8A:26pp, 8.84MB; http://palaeo-electronica.org/2010_2/198/index.html
Mallison, H. 2010b. The digital Plateosaurus II: an assessment of the range of motion of the limbs and vertebral column and of previous reconstructions using a digital skeletal mount. Acta Palaeontologica Polonica, 55:433-458. doi:10.4202/app.2009.0075.
Mallison, H. 2010c. CAD assessment of the posture and range of motion of Kentrosaurus aethiopicus Hennig 1915. Swiss Journal of Geosciences, 103:211-233. doi:10.1007/s00015-010-0024-2.
Mallison, H. 2011a. Plateosaurus in 3D: How CAD models and kinetic/dynamic modeling help bringing an extinct animal to life, p. 219-236. In Klein, N., Remes, K., Gee, C., and Sander, P.M. (eds.), Biology of the Sauropod Dinosaurs: Understanding the life of giants. Life of the Past. Indiana University Press, Bloomington.
Mallison, H. 2011b. Rearing Giants – kinetic/dynamic modeling of sauropod bipedal and tripodal poses, p. 237-250. In Klein, N., Remes, K., Gee, C.T., and Sander, P.M. (eds.), Biology of the Sauropod Dinosaurs: Understanding the life of giants. Life of the Past. Indiana University Press, Bloomington.
Martínez, R.N. 2009. Adeopapposaurus mognai, gen. et sp. nov (Dinosauria: Sauropodomorpha), with comments on adaptations of basal sauropodomorpha. Journal of Vertebrate Paleontology, 29:142-164.
Moser, M. 2003. Plateosaurus engelhardti Meyer, 1837 (Dinosauria, Sauropodomorpha) aus dem Feuerletten (Mittelkeuper; Obertrias) von Bayern. Zitteliana Reihe B, Abhandlungen der Bayerischen Staatssammlung fuer Palaeontologie und Geologie, 24:1-186.
Norman, D.B. 2004. Basal Iguanodontia, p. 413-437. In Weishampel, D.B., Dodson, P., and Osmolska, H. (eds.), The Dinosauria, 2nd Edition. University of California Press, Berkeley.
Norman, D.B., Sues, H.-D., Witmer, L.M., and Coria, R.A. 2004. Basal Ornithopoda, p. 393-412. In Weishampel, D.B., Dodson, P., and Osmolska, H. (eds.), The Dinosauria, 2nd Edition. University of California Press, Berkeley.
Ostrom, J.H. 1970. Stratigraphy and paleontology of the Cloverly Formation (Lower Creataceous) of the Bighorn Basin area, Wyoming and Montana. Bulletin of the Peabody Museum of Natural History, 35:1-234.
Paul, G.S. 1987. The science and art of restoring the life appearance of dinosaurs and their relatives. A rigorous how-to guide, p. 5-49. In Czerkas, S.J. and Olson, E.C. (eds.), Dinosaurs Past and Present Volume II. Natural History Museum of Los Angeles County Press, Los Angeles.
Paul, G.S. 1997. Dinosaur models: the good, the bad, and using them to estimate the mass of dinosaurs, p. 129-154. In Wolberg, D.L., Stump, E., and Rosenberg, G. (eds.), Dinofest International: Proceedings of a Symposium held at Arizona State University. Academy of Natural Sciences, Philadelphia.
Paul, G.S. 2000. Restoring the life appearance of dinosaurs, p. 78-106. In Paul, G.S. (ed.), The Scientific American Book of Dinosaurs. Byron Press and Scientific American, New York.
Prieto-Márques, A. and Norell, M.A. 2011. Redescription of a nearly complete skull of Plateosaurus (Dinosauria: Sauropodomorpha) from the Late Triassic of Trossingen (Germany). American Museum Novitates, 3727:1-58.
Rauhut, O.W.M., Fechner, R., Remes, K., and Reis, K. 2011. How to get big in the Mesozoic: the evolution of the sauropodomorph body plan, p. 119-149. In Klein, N., Remes, K., Gee, C.T., and Sander, P.M. (eds.), Biology of the Sauropod Dinosaurs: Understanding the life of giants. Indiana University Press, Bloomington.
Reisz, R.R., Scott, D., Sues, H.-D., Evans, D.C., and Raath, M.A. 2005. Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance. Science, 309:761-764.
Rowe, T.B., Sues, H.-D., and Reisz, R.R. 2010. Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon. Proceedings of the Royal Society B: Biological Sciences, 278:1044-1053.
Sander, P.M. 1992. The Norian Plateosaurus bonebeds of central Europe and their taphonomy. Palaeogeography, Palaeoclimatology, Palaeoecology, 93:255-296.
Sander, P.M. and Klein, N. 2005. Developmental plasticity in the lift history of a prosauropod dinosaur. Science, 310:1800-1802.
Santa-Luca, A.P. 1980. The postcranial skeleton of Heterodontosaurus tucki (Reptilia, Ornithischia) from the Stormberg of South Africa. Annals of the South African Museum, 79:159-211.
Schwarz, D., Wings, O., and Meyer, C.A. 2007. Super sizing the giants: first cartilage preservation at a sauropod dinosaur limb joint. Journal of the Geological Society London, 164:61-65.
Seebacher, F. 2001. A new method to calculate allometric length-mass relationships of dinosaurs. Journal of Vertebrate Paleontology, 21:51-60.
Sertich, J.J.W. and Loewen, M.A. 2010. A new basal sauropodomorph dinosaur from the Lower Jurassic Navajo Sandstone of Southern Utah. PLoS ONE, 5(3):e9789. doi:10.1371/journal.pone.0009789.
Tornier, G. 1912. [no title]. Zeitschrift der Deutschen Geologischen Gesellschaft, Monatsberichte, 64:2-24.
Upchurch, P., Barrett, P.M., and Galton, P.M. 2007. A phylogenetic analysis of basal sauropodomorph relationships: implications for the origin of sauropod dinosaurs, p. 57-90. In Barrett, P.M. and Batten, D.J. (eds.), Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (Special Papers in Palaeontology 77). Blackwell Scientific Publications, Oxford.
Van Heerden, J. 1997. Prosauropods, p. 242-263. In Farlow, J. and Brett-Surman (eds.), The complete dinosaur. Indiana University Press, Bloomington.
von Huene, F. 1905. Über die Trias-Dinosaurier Europas. Zeitschrift der Deutschen Geologischen Gesellschaft, 57:345-349.
von Huene, F. 1926. Vollständige Osteologie eines Plateosauriden aus dem schwäbischen Keuper. Geologische und Palaeontologische Abhandlungen, Neue Folge, 15:139-179.
von Huene, F. 1932. Die fossile Reptile- Ordnung Saurischia, ihre Entwicklung und Geschichte, Teil I (Text). Monographien zur Geologie und Palaeontologie, Series 1, 4:1-361.
von Meyer, H. 1837. Mitteilung an Prof. Bronn. Neues Jahrbuch für Geologie und Paläontologie, 1837: 316.
Weishampel, D.B. 1986. Trossingen: E. Fraas, F. von Huene, R. Seemann, and the "Schwäbische Lindwurm" Plateosaurus, p. 249-253. In Reif, W.-E. and Westphal, F. (eds.), Third Symposium on Terrestrial Ecosystems, Short Papers. Attempto, Tübingen.
Weishampel, D.B. and Westphal, F. 1986. Die Plateosaurier von Trossingen im Geologischen Institut der Eberhard-Karls-Universität Tübingen. Ausstellungskataloge der Universität Tübingen, 19:1-27.
Wellnhofer, P. 1994. Prosauropod dinosaurs from the Feuerletten (Middle Norian) of Ellingen near Weissenburg in Bavaria. Revue de Paléobiologie, Volume special, 7:263-271.
Yates, A.M. 2003a. Species taxonomy of the sauropodomorph dinosaurs from the Löwenstein Formation (Norian, Late Triassic) of Germany. Palaeontology, 46:317-337.
Yates, A.M. 2003b. A new species of the primitive dinosaur Thecodontosaurus (Saurischia: Sauropodomorpha) and its implications for the systematics of early dinosaurs. Journal of Systematic Palaeontology, 1:1-42.
Yates, A.M. 2007. The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria), p. 9-55. In Barret, P.M. and Batten, D.J. (eds.), Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs (Special Papers in Palaeontology 77). Blackwell Scientific Publications, Oxford.
Yates, A.M. and Kitching, J.W. 2003. The earliest known sauropod dinosaur and the first steps towards sauropod locomotion. Proceedings of the Royal Society B: Biological Sciences, 270:1753-1758.
