 Geochronology of the Upper Alturas Formation, northern California: Implications for the Hemphillian-Blancan North American Land Mammal Age boundary
Geochronology of the Upper Alturas Formation, northern California: Implications for the Hemphillian-Blancan North American Land Mammal Age boundary
Article number: 17.3.43A
https://doi.org/10.26879/493
Copyright Palaeontological Association, November 2014
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 14 May 2014. Acceptance: 12 November 2014
{flike id=985}
ABSTRACT
Fossil vertebrates from the Alturas Formation in northern California have previously been considered important for defining the age of the boundary between the Hemphillian and Blancan North American Land Mammal Ages. Diatomaceous mudstone of the upper Alturas Formation contain fossil mammals including the arvicoline rodent Mimomys (Ogmodontomys) sawrockensis that is diagnostic of Blancan faunas. New paleomagnetic and geochemical data from the upper Alturas Formation constrain the age of the first stratigraphic occurrence of M. (O.) sawrockensis at Crowder Flat Road to between 4.5 and 4.6 Ma. This age is approximately 0.2-0.4 Ma younger than previously reported such that the oldest record of Mimomys in North America, south of 55oN, is from Panaca, Nevada, and is constrained geochronologically to be approximately 4.9 Ma. The Hemphillian - Blancan North American Land Mammal Age boundary probably occurs within magnetic polarity Chron C3n.3r at approximately 4.9 Ma.
Steven R. May. University of Texas, Jackson School of Geosciences, Vertebrate Paleontology Laboratory, R7600, Austin, Texas 78758, USA. srmay@utexas.edu
Andrei M. Sarna-Wojcicki. United States Geological Survey, 345 Middlefield Road MS-973, Menlo Park, California 94025, USA. asarna@usgs.gov
Everett H. Lindsay. University of Arizona, Department of Geosciences, Gould-Simpson Building #77, 1040 E 4th St., Tucson, California 85721, USA. ehlind@cox.net
Michael O. Woodburne. Museum of Northern Arizona, 3101 N. Ft. Valley Rd., Flagstaff, California 86001, USA. mikew@npgcable.com
Neil D. Opdyke. University of Florida, Dept. of Geological Sciences, Gainesville, FL 32611. drno@ufl.edu
Elmira Wan. United States Geological Survey, 345 Middlefield Road, Menlo Park, California 94025, USA. ewan@usgs.gov
David B. Wahl. United States Geological Survey, 345 Middlefield Road, Menlo Park, California 94025, USA. dwahl@usgs.gov
Holly Olson. United States Geological Survey, 345 Middlefield Road, Menlo Park, California 94025, USA. olsonholly@hotmail.com
Keywords: geochronology; Hemphillian; Blancan; arvicoline
Final citation: May, Steven R., Sarna-Wojcicki, Andrei M., Lindsay, Everett H., Woodburne, Michael O., Opdyke, Neil D., Wan, Elmira, Wahl, David B., and Olson, Holly. 2014. Geochronology of the upper Alturas Formation, northern California: Implications for the Hemphillian-Blancan North American Land Mammal Age boundary. Palaeontologia Electronica 17.3.43A: 1-13. https://doi.org/10.26879/493
palaeo-electronica.org/content/2014/985-alturas-geochronology
INTRODUCTION
North American land mammal ages (Wood et al., 1941) have been classically viewed as biochronologic units based on the evolution and dispersal of fossil mammals. Their utility for correlation is dependent on the ability to define unambigous boundaries and to characterize the intervening time intervals by the joint occurrence of multiple taxa (Woodburne, 2004). Widespread immigration events of single or multiple taxa are commonly regarded as potential criteria for boundary definition (Repenning, 1967; Woodburne, 1996). Historically, fossil mammals from the Alturas Formation in California have played a key role in the definition of the base of the Blancan mammal age based on the first occurrence of the arvicoline rodent Mimomys and interpretation of associated geochronologic data.
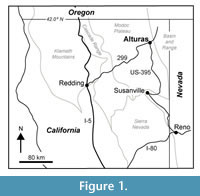 The Alturas Formation is exposed in valleys of the Modoc Plateau in Northeastern California and includes approximately 200-300 m of volcaniclastic and diatomaceous sedimentary rocks that were deposited during the Late Miocene and Pliocene (Figure 1). Strata of the Alturas Formation are generally flat lying and are commonly overlain by olivine basalts. Collins (1999) measured ~118 m of Alturas Formation sediments at Rattlesnake Butte and ~170 meters of Alturas Formation at Crowder Flat Road (Figure 2). She recognized six different depositional facies represented by pyroclastic/debris flows, fluvial, lacustrine, and pyroclastic / hydrovolcanic strata and interpreted these strata to record deposition in a continental back-arc setting. Especially common in the upper 50 meters of the Crowder Flat Road section, Collins also documented the presence of diatomite and diatomaceous mudstone representing a well-developed lacustrine environment. Krebs et al. (1987) described a Pliocene diatom assemblage from the Alturas Formation, presumably from the Crowder Flats Road section based on reference to an “in press” paper by C.A. Repenning.
The Alturas Formation is exposed in valleys of the Modoc Plateau in Northeastern California and includes approximately 200-300 m of volcaniclastic and diatomaceous sedimentary rocks that were deposited during the Late Miocene and Pliocene (Figure 1). Strata of the Alturas Formation are generally flat lying and are commonly overlain by olivine basalts. Collins (1999) measured ~118 m of Alturas Formation sediments at Rattlesnake Butte and ~170 meters of Alturas Formation at Crowder Flat Road (Figure 2). She recognized six different depositional facies represented by pyroclastic/debris flows, fluvial, lacustrine, and pyroclastic / hydrovolcanic strata and interpreted these strata to record deposition in a continental back-arc setting. Especially common in the upper 50 meters of the Crowder Flat Road section, Collins also documented the presence of diatomite and diatomaceous mudstone representing a well-developed lacustrine environment. Krebs et al. (1987) described a Pliocene diatom assemblage from the Alturas Formation, presumably from the Crowder Flats Road section based on reference to an “in press” paper by C.A. Repenning.
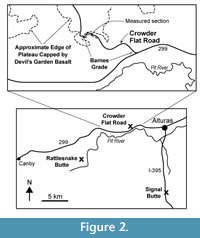 The Alturas Formation is overlain by Pliocene olivine basalts that have been described by a number of authors including McKee et al. (1983) and Carmichael et al. (2006). Previously referred to as the Warner basalt, Carmichael et al. (2006) described the basalt at the top of the Barnes Grade along the Crowder Flat Road section as the Devil’s Garden Basalt and published an 40Ar/39Ar plateau age of 4.31 +/- 0.18 Ma.
The Alturas Formation is overlain by Pliocene olivine basalts that have been described by a number of authors including McKee et al. (1983) and Carmichael et al. (2006). Previously referred to as the Warner basalt, Carmichael et al. (2006) described the basalt at the top of the Barnes Grade along the Crowder Flat Road section as the Devil’s Garden Basalt and published an 40Ar/39Ar plateau age of 4.31 +/- 0.18 Ma.
Although the original collection of fossil mammals from the Crowder Flats Road section was made by J.A. Shotwell in 1958 (UO 2424), plant and animal fossils from the Alturas Formation had been known for many decades. Dorf (1933) was the first to use the name Alturas Formation in his discussion of fossil floras from California and considered it to be “upper Pliocene” on the basis of stratigraphic relations and vertebrate fossils whose age assignment were referenced to Chester Stock, oral communication, 1929. LaMotte (1936) continued the use of the name Alturas Formation and described it as consisting of isolated exposures of conglomerate, sandstone, ash, and shale and referenced localities at Rattlesnake Butte, Davis Creek, Tuledad Canyon, and Warner Valley. R.A. Stirton contributed a short section of LaMotte’s 1936 paper wherein he identified Vulpes sp., Rhinoceratidae, Tayassuidae, Neohipparion sp., Alticamelus sp., Mastodontinae, Camelidae, Felis sp., and Carnivora from six localities in “the vicinity of Alturas,” presumably from the Alturas formation at Rattlesnake Butte according to LaMotte. Stirton concluded “there is not sufficient identifiable material in our collection from Alturas for an accurate determination of the age of the stratum from which they were obtained.” The general character of this fauna and a radiometric date from near Rattlesnake Butte (Evernden et al., 1964) suggested a Hemphillian age.
Axelrod (1944) provided further discussion of the fossil flora from the Alturas Formation based on re-examination of previous materials and from additional material obtained from the “Alturas formation near Rattlesnake Butte.” His fossil locality (UCMP 117) is described as being 1/2 mile SW of Rattlesnake Butte along the north bank of the Pit River (Figure 2). He describes the Alturas Formation as including a few hundred feet of relatively horizontal strata including tuffaceous sandstones, conglomerates, tuffaceous shales, and clays. Axelrod (1944) concluded a Pliocene age for the Alturas flora. Evernden et al. (1964) published a K/Ar date from plagioclase in a tuff near the base of the Alturas Formation of 8.1 Ma.
The first stratigraphic occurrence of the microtine rodent Mimomys (Ogmodontomys) sawrockensis in the Alturas Formation was interpreted by Repenning (1987, 2003) to be dated at ~ 4.8 Ma and was considered by him to be the oldest occurrence in North America and therefore to constrain the base of the Blancan Land Mammal Age at approximately 4.8 Ma. In their review of the Blancan Land Mammal Age, Bell et al. (2004) stated that “the Blancan is currently defined by the first appearance in North America south of 55o N latitude of arvicoline rodents in the genera Mimomys, Ogmodontomys, and Ophiomys.” They refered to Repenning’s (1987) discussion of Mimomys (Ogmodontomys) sawrockensis from Alturas as the earliest occurrence of these taxa, but also noted a potentially earlier occurrence of Mimomys from Panaca, Nevada (Lindsay et al., 2002).
The objective of this study is to provide new data and reevaluate existing data concerning the age of the upper Alturas Fauna and to review the significance of this age with respect to the Hemphillian-Blancan boundary.
Terminology
We utilize the notation of “m” and “M” for lower and upper molars, respectively. Abbreviations associated with fossil localities include: UCMP - University of California Museum of Paleontology, UO - University of Oregon, and USGS - U.S. Geological Survey.
METHODS
Tephrochronology
Volcanic glasses separated from all tephra samples were analyzed by electron-microprobe to measure the abundances of Na, Si, Al, Fe, Mg, Mn, Ca, Ti, and K. Detailed laboratory procedures and methods of chemical analysis are described in Sarna-Wojcicki et al. (2005, 2011). Chemical analysis of the volcanic glasses was performed by electron microprobe. We used GSC and An40 as standards, and RLS 132, a homogenous obsidian from La Puebla, Mexico, as an internal standard (Myers et al., 1976). The ZAF data reduction program was used to obtain oxide concentrations. Internal, polished surfaces of 15 to 20 individual glass shards were analyzed for each sample. The means of the individual shard analyses are averaged for the oxides to obtain an overall composition for each sample.
Results of analyses of the volcanic glass from the samples collected in this study were compared with ~5800 analyses of tephra samples contained in the U.S. Geological Survey’s database of Upper Neogene tuffs collected from the western U.S., Alaska, and Mexico. These data are stored at the U.S. Geological Survey's Tephrochronology Laboratory in Menlo Park, California. Analytical data on the chemical composition of volcanic glasses of tephra layers that we obtained in this study were first evaluated using the similarity coefficient of Borchardt et al. (1972), and Borchardt (1974). The similarity coefficient is used as a guide to select a pool of candidate samples that are further evaluated in terms of the chemical and geological criteria for the closeness of a match. The means of oxide analyses for the individual shards are averaged to obtain an overall composition for each sample. We used a computer program that compares any single sample with all previously analyzed samples, for those elements that are considered the most reliable in chemical identification of tephra layers, and that allow us to distinguish most clearly between tephra layers of similar composition (Sarna-Wojcicki et al., 1984; Sarna-Wojcicki and Davis, 1991; Sarna-Wojcicki, 2000). This approach lists the tephra samples that match most closely to the sample that is being evaluated for a specified set of elements, and these samples are ranked in order of the value of the similarity coefficient. The highest ranking samples represent a pool of candidates for further evaluation for correlation.
For a more complete and detailed discussion of field criteria, petrographic characteristics, mineralogy, chemical analysis of glass, and data evaluation methods used here, see Sarna-Wojcicki (1971), Sarna-Wojcicki et al. (1984), Sarna-Wojcicki and Davis (1991), Sarna-Wojcicki (2000), and Sarna-Wojcicki et al. (2005, 2011).
Paleomagnetism
Six to eight oriented hand samples were collected by May and Repenning in 1984 from each of 10 initial sites along Crowder Flat Road. A site consisted of a single sedimentary bed, and samples were separated laterally by no more than 2 meters. This initial sampling spanned approximately 35 meters of the Alturas Formation below the contact with the Devil’s Garden Basalt. A single oriented block of the basalt was also collected, and multiple 1-inch diameter cores were drilled from this block. Bulk sedimentary rock samples were also collected at four of the sites for analysis of magnetic mineralogy.
 Oriented hand samples were cut to fit 1 in3 plastic boxes and all paleomagnetic and rock magnetic measurements from this initial sample set were made at the University of Arizona Department of Geosciences paleomagnetic laboratory. Both thermal and alternating field demagnetization was applied to individual samples to determine the most effective procedure for defining the primary remanant magnetization.
Oriented hand samples were cut to fit 1 in3 plastic boxes and all paleomagnetic and rock magnetic measurements from this initial sample set were made at the University of Arizona Department of Geosciences paleomagnetic laboratory. Both thermal and alternating field demagnetization was applied to individual samples to determine the most effective procedure for defining the primary remanant magnetization.
Demagnetization experiments for the initial set of samples included stepwise alternating field demagnetization at 5 mT intervals up to 30 mT on all specimens. Higher demagnetization steps up to 70 mT were performed on selected specimens. AF demagnetization up to 30 mT generally resulted in a loss of intensity from 30% to 95% of the NRM. Because of a viscous normal component present in most of the primary reversed samples, an initial increase in intensity was often observed. Stepwise thermal demagnetization was applied to several samples in steps from 90 to 600°C using a modified ASC TD-48 thermal specimen demagnetizer. Stable magnetizations were defined using both AF and thermal demagnetization (Figure 3).
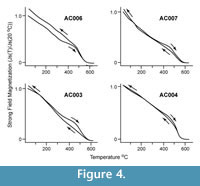 Jsat-T experiments on magnetic extracts from bulk sedimentary rock samples at sites AC003, AC004, AC006, and AC007 were performed to help identify the primary magnetic phase. A strong field thermomagnetic balance was used to measure the apparent weight change of the sample in an applied field as the temperature was increased then decreased. All experiments were carried out in < 1 atmosphere of argon with a heating rate of 15°C/minute. Jsat-T curves observed in the thermomagnetic experiments are illustrated in Figure 4. These experiments demonstrated a general loss in magnetic intensity as a function of heating to 610oC with all remanance removed by 580oC. This suggests that low Ti-magnetite is the primary magnetic mineral consistent with the results of the AF and thermal demagnetization experiments. We interpret the magnetization in the Alturas Formation sediments to be a primary detrital remanant magnetization acquired at the time of deposition.
Jsat-T experiments on magnetic extracts from bulk sedimentary rock samples at sites AC003, AC004, AC006, and AC007 were performed to help identify the primary magnetic phase. A strong field thermomagnetic balance was used to measure the apparent weight change of the sample in an applied field as the temperature was increased then decreased. All experiments were carried out in < 1 atmosphere of argon with a heating rate of 15°C/minute. Jsat-T curves observed in the thermomagnetic experiments are illustrated in Figure 4. These experiments demonstrated a general loss in magnetic intensity as a function of heating to 610oC with all remanance removed by 580oC. This suggests that low Ti-magnetite is the primary magnetic mineral consistent with the results of the AF and thermal demagnetization experiments. We interpret the magnetization in the Alturas Formation sediments to be a primary detrital remanant magnetization acquired at the time of deposition.
 Nine additional sites (011- 019) were collected in 2008 to extend the magnetic polarity stratigraphy farther down section along the Crowder Flat Road and to provide additional resolution elsewhere in the section. One site (001) was collected from the light gray ash in a roadcut along US 395 at Signal Butte, south of Alturas (Figure 2). During this field work, five samples were collected at each site, and samples were again placed in small plastic cubes. These samples were processed at the University of Florida in 2008 using AF demagnetization in progressive steps from 0 - 80 mT. Interpretation of magnetic polarity for this second set of samples was unambiguous and the results are consistent with the initial samples collected in 1984.
Nine additional sites (011- 019) were collected in 2008 to extend the magnetic polarity stratigraphy farther down section along the Crowder Flat Road and to provide additional resolution elsewhere in the section. One site (001) was collected from the light gray ash in a roadcut along US 395 at Signal Butte, south of Alturas (Figure 2). During this field work, five samples were collected at each site, and samples were again placed in small plastic cubes. These samples were processed at the University of Florida in 2008 using AF demagnetization in progressive steps from 0 - 80 mT. Interpretation of magnetic polarity for this second set of samples was unambiguous and the results are consistent with the initial samples collected in 1984.
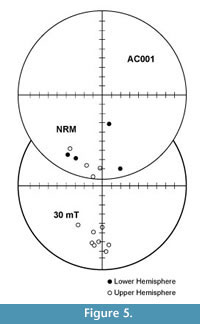
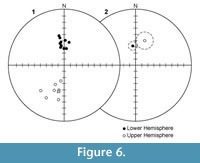
Progressive demagnetization data from all specimens were analyzed with vector demagnetization diagrams. Within each site, all samples record the same polarity, but the dispersion was occasionally quite high. Within-site alpha95 values ranged from a few degrees to >20o. Polarity determinations for all sites were unambiguous based on the position and trend of the horizontal and vertical components as seen in equal area stereo plots of the data (Figure 5). The site mean directions for sites with alpha95 < 20o barely pass a reversals test at the 95% confidence interval (Figure 6). This is likely due to a present field secondary magnetization that was not completely removed from the reversed polarity sites. No fold test was available as all strata within the Crowder Flat Road section are essentially flat lying.
RESULTS
 Tephrochronology
Tephrochronology
Results of chemical analysis of volcanic glass from the tephra layers are presented in Table 1. Most of the 13 layers from this section do not match other, previously analyzed tephra layers in the USGS data base, and we suspect that the former were derived from the nearby Cascade Range to the west or from other local sources just east of the Cascades. One of these layers, however, matches well with widespread units that have been previously identified and dated elsewhere in the western conterminous U.S.
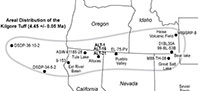
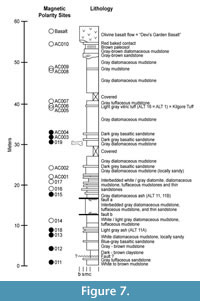
On the basis of the chemical composition of its volcanic glass, we identify a light-gray vitric tuff, situated 15.6 m below the Devil’s Garden Basalt, as the Kilgore Tuff, dated 4.45 ± 0.05 Ma (Morgan and McIntosh, 2005) (Table 1, ALT-1, ALT-16) (Figure 7). The type locality of the Kilgore Tuff, just south of the Snake River Plain in Idaho, exhibits reversed magnetic polarity and represents the youngest, large-volume ignimbrite and plinian pumice fall erupted from the Heise caldera (Figure 8). This caldera and its associated deposits are the immediate predecessor of the Yellowstone caldera situated to the east of the Heise volcanic field. Geochemical analyses of the volcanic glass from this tuff, and its correlative samples in the Alturas area (ALT-1, ALT-16, ALT-2A-N) match well (Table 1).
 In the area of this study, the Kilgore Tuff is also present in a road cut along U.S. Highway 395 at Signal Butte, about 7 miles south of Alturas, California (Figure 2). Chemical analysis of the tuff indicate that most of the shards present in the tephra layer at Signal Butte match well with the Kilgore Tuff, but a small number of them are chemically different, representing a minor mode, possibly owing to detrital contamination with a different ash. Paleomagnetic sites from the Kilgore Tuff at Crowder Flat Road (AC006) and Signal Buttes (001) both exhibit reversed polarity as reported in the type region (Morgan and McIntosh, 2005).
In the area of this study, the Kilgore Tuff is also present in a road cut along U.S. Highway 395 at Signal Butte, about 7 miles south of Alturas, California (Figure 2). Chemical analysis of the tuff indicate that most of the shards present in the tephra layer at Signal Butte match well with the Kilgore Tuff, but a small number of them are chemically different, representing a minor mode, possibly owing to detrital contamination with a different ash. Paleomagnetic sites from the Kilgore Tuff at Crowder Flat Road (AC006) and Signal Buttes (001) both exhibit reversed polarity as reported in the type region (Morgan and McIntosh, 2005).
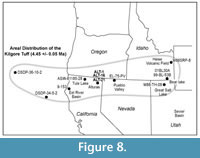 In addition to the Alturas area, the Kilgore Tuff has been identified at several other locations in the western conterminous U.S., interbedded with non-marine sediments, as well as in marine sediments of the northeastern Pacific Ocean (Figure 8; Table 1). The presence of the Kilgore Tuff in marine sediments in deep-ocean cores of the northeast Pacific Ocean presents the possibility of comparing closely contemporaneous marine and non-marine fauna and flora, and interpreting changes in non-marine faunas such as those discussed here within a broader paleoclimatic context.
In addition to the Alturas area, the Kilgore Tuff has been identified at several other locations in the western conterminous U.S., interbedded with non-marine sediments, as well as in marine sediments of the northeastern Pacific Ocean (Figure 8; Table 1). The presence of the Kilgore Tuff in marine sediments in deep-ocean cores of the northeast Pacific Ocean presents the possibility of comparing closely contemporaneous marine and non-marine fauna and flora, and interpreting changes in non-marine faunas such as those discussed here within a broader paleoclimatic context.
Magnetic Polarity Stratigraphy
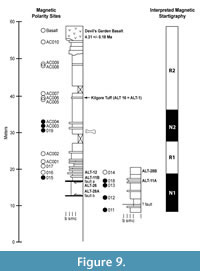 The resulting magnetic polarity stratigraphy is shown in Figure 9. The upper part of the section including the Devil’s Garden Basalt is reversed polarity down though site AC005 (R2). This reversed polarity interval includes site AC006, which is the Kilgore Tuff. A normal polarity interval is then observed from site AC004 through 019 (N2). This normal polarity interval includes the primary fossil mammal locality (UO 2424 = VF 155) that records the first stratigraphic occurrence of Mimomys (Ogmodontomys) sawrockensis. Another reversed polarity interval is recorded in sites AC002 through 016 (R1). Normal polarity is then observed again at site 015 just above the first normal fault below the basalt (fault a) (N1). A reversed to normal transition is recorded again below the second fault (fault b) between sites 014 and 018. Based on our analysis of the throw on these two faults, we interpret this to be the same polarity transition as observed between sites 016 and 015. This correlation implies that 015 is roughly equivalent to 018. Although the equivalent tuffs sampled at ALT-11B and ALT-11A are geochemically distinct, they are so similar that we believe they represent slightly different age eruptions from a single evolving magma chamber. Normal polarity continues down section through site 011. A fault may be present between sites 012 and 011, but the displacement on this fault is uncertain.
The resulting magnetic polarity stratigraphy is shown in Figure 9. The upper part of the section including the Devil’s Garden Basalt is reversed polarity down though site AC005 (R2). This reversed polarity interval includes site AC006, which is the Kilgore Tuff. A normal polarity interval is then observed from site AC004 through 019 (N2). This normal polarity interval includes the primary fossil mammal locality (UO 2424 = VF 155) that records the first stratigraphic occurrence of Mimomys (Ogmodontomys) sawrockensis. Another reversed polarity interval is recorded in sites AC002 through 016 (R1). Normal polarity is then observed again at site 015 just above the first normal fault below the basalt (fault a) (N1). A reversed to normal transition is recorded again below the second fault (fault b) between sites 014 and 018. Based on our analysis of the throw on these two faults, we interpret this to be the same polarity transition as observed between sites 016 and 015. This correlation implies that 015 is roughly equivalent to 018. Although the equivalent tuffs sampled at ALT-11B and ALT-11A are geochemically distinct, they are so similar that we believe they represent slightly different age eruptions from a single evolving magma chamber. Normal polarity continues down section through site 011. A fault may be present between sites 012 and 011, but the displacement on this fault is uncertain.
A silver-gray ash is correlated geochemically across the two faults (samples ALT-12, ALT-26; ALT-28A; and ALT-28B are all geochemically indistinguishable) (Figure 9). This correlation is consistent with a throw of ~2.4 m across fault a and 0.5 m across fault b. Correlation across faults a and b results in the magnetic polarity stratigraphy as shown in Figure 10.
Correlation of the magnetic polarity stratigraphy with the magnetic polarity time scale of Gradstein et al. (2012) is straightforward given the geochronologic control from the Devil’s Garden Basalt and the Kilgore Tuff. We interpret the stratigraphically lowest normal polarity magnetozone (N1) as C3n.3n. The next younger normal magnetozone (N2) is interpreted as C3n.2n. R2 includes the Kilgore Tuff dated at 4.45 +/- 0.05 Ma (Morgan and McIntosh, 2005) and the Devils Garden Basalt dated at 4.31 +/- 0.18 Ma (Carmichael et al., 2006). This polarity zonation and the associated radiometric dates are consistent with the time scale of Gradstein et al. (2012).
DISCUSSION AND CONCLUSIONS
 In Repenning’s (2003) review of Mimomys in North America, he discussed M. sawrockensis from the Upper Alturas fauna and assigned it an age of 4.8 Ma. At the time, he interpreted this to be the earliest occurrence of Mimomys in North America and defined the beginning of the Blancan North American Land Mammal Age. Repenning stated: “The immigration of Mimomys to North America marks the beginning of the Blancan Mammal Age by original definition. The earliest dated North American record is the Upper Alturas Fauna of California, which is well dated by paleomagnetic stratigraphy, tephra “fingerprinting”, and potassium-argon dating at 4.8 Ma.” This reflected an interpretation based on a previous K-Ar date from the Devil’s Garden Basalt of 4.7 +/ 0.5 Ma (cited by Repenning, 2003 as “Silberman pers. comm.”) and an associated interpretation that the normal polarity zone that included the first stratigraphic occurrence of Mimomys was Chron C3n.4n.
In Repenning’s (2003) review of Mimomys in North America, he discussed M. sawrockensis from the Upper Alturas fauna and assigned it an age of 4.8 Ma. At the time, he interpreted this to be the earliest occurrence of Mimomys in North America and defined the beginning of the Blancan North American Land Mammal Age. Repenning stated: “The immigration of Mimomys to North America marks the beginning of the Blancan Mammal Age by original definition. The earliest dated North American record is the Upper Alturas Fauna of California, which is well dated by paleomagnetic stratigraphy, tephra “fingerprinting”, and potassium-argon dating at 4.8 Ma.” This reflected an interpretation based on a previous K-Ar date from the Devil’s Garden Basalt of 4.7 +/ 0.5 Ma (cited by Repenning, 2003 as “Silberman pers. comm.”) and an associated interpretation that the normal polarity zone that included the first stratigraphic occurrence of Mimomys was Chron C3n.4n.
Repenning (2003) described the primary fossil locality for M. sawrockensis in the Alturas Formation as:
“Upper Alturas fauna: Barnes Grade, Modoc County, California; USGS, VPR M-1505 (transferred to the Denver office in 1985) (= CAS locality 36805 collected for diatoms by G.D. Hanna, C. Chesterman, and C. Jennings in Sept. 1959; = UO locality 26916 collected by J.A. Shotwell in the late 1950’s). Along Barnes Grade of Crowder Flat road leading up to Big Sage Reservoir, north of US 299 and 4 mi west of Alturas, California about 72 ft below a capping basalt (dated 4.7 +/- 0.5 Ma by M.L. Silberman, personal commun. ..., 6 ft below prominent white, diatomaceous tephra deposit dated 4.8 Ma by “fingerprint” correlation to a marine biochronology core off the California Coast (A.M. Sarna-Wojcicki, personal commun., 1985), and 5 ft above the Thvera event of the Gilbert Chron in a paleomagnetic section measured up Barnes Grade to the capping basalt by S.R. May and myself in 1984... This was the original discovery locality of the Upper Alturas fauna.”
In addition to the new geochronological data presented here, it should be noted that “UO 26916” is a specimen number rather than a University of Oregon locality number. The actual locality is UO 2424 (= VF 155) (T. Fremd, personal commun., 2009) and is presumed to be equivalent to USGS, VPR M-1505. The collection is still housed at the University of Oregon and includes 6 specimens of Mimomys (T. Fremd, personal commun., 2009).
The new data and interpretations presented in this paper bear directly on the age of this locality described in Repenning (2003). The Ar/Ar data published by Carmichael et al. (2006) for the basalt at the top of the Crowder Flat road section indicate an age of 4.31 +/- 0.18 Ma rather that 4.7 Ma. The ash that occurs above the fossil mammal locality is now known to be the Kilgore Tuff that has been radiometrically dated at 4.45 +/- 0.05 Ma (Morgan and McIntosh, 2005). The normal polarity interval associated with the fossil mammal locality is now interpreted to be C3n.2n (4.49-4.63 Ma, Gradstein et al., 2012). The fossil mammal locality along Crowder Flat road that contains the first stratigraphic occurrence of M. sawrockensis is now interpreted to be approximately 4.5 - 4.6 Ma and therefore not the first occurrence of Mimomys in North America.
The geochronology presented here applies to only the upper 55 meters of the Alturas Formation while Collins (1999) measured approximately 170 meters of total section along Crowder Flat road. Older strata within this section are almost certainly Hemphillian in age. Repenning (2003) suggested there were two distinct faunas from the Alturas Formation with his use of “Upper Alturas Fauna” as opposed to the Hemphillian age fauna that he refers to through citations of Axelrod (1944), Evernden et al. (1964) and Wood et al. (1941). Therefore, the Hemphillian - Blancan boundary is likely captured within the Alturas Formation although the exact location of the boundary is not yet defined in a single, continuous, fossiliferous section.
In their review of the Hemphillian-Blancan boundary, Bell et al. (2004) state that “the Blancan North American Land Mammal Age is defined by the first appearance in North America, south of 55o N, of arvicoline rodents in the genera Mimomys, Ogmodontomys, and Ophiomys. ” Given the new age constraints presented in this paper for the first occurrence of Mimomys in the Alturas Formation, the oldest occurrence of Mimomys in North America is interpreted to be at Panaca, Nevada (Lindsay et al., 2002). Mou (1997) named Mimomys panacaensis from the Panaca Formation as a new microtine species that is similar to M. mcknighti and M. sawrockensis and occurs first within strata correlated with chron C3n.3r. Mou (1998) further argued that the enamel microstructure of M. panacaensis differs from Ogmodontomys and Ophiomys, but is similar to Eurasian Mimomys. Mou (2011) named a new genus of arvicoline rodent from Panaca as Nevadomys, but considered that whereas M. panacaensis was an immigrant, Nevadomys was endemic to North America.
The Upper Alturas fauna is significantly younger than the Panaca fauna and only constrains the Hempillian-Blancan boundary to be older than ~4.6 Ma. Lindsay et al. (2002) concluded that paleontologic, radioisotopic, and magnetostratigraphic data from Panaca were consistent with placement of the Hemphillian/Blancan boundary near the boundary between chrons C3n.3r and C3n.4n. Magneostratigraphic and tephrochronologic data from the Horned Toad Formation in the northern Mojave Desert, California, indicate that the latest Hemphillian Warren fauna is also correlative with chron C3n.3r (May et al., 2011). This suggests that the Hemphillian - Blancan North American Land Mammal Age boundary occurs within C3n.3r or approximately 4.9 Ma as also concluded by Gradstein et al. (2012).
ACKNOWLEDGMENTS
We would like to acknowledge C. Repenning for introducing us to the Crowder Flat Road section of the Alturas Formation and for pointing out its potential significance for the Hemphillian-Blancan boundary through numerous discussions. T. Fremd kindly provided help with the University of Oregon mammalian paleontology collection. The manuscript was improved by comments from Dr. L. Jacobs and one anonymous reviewer.
REFERENCES
Axelrod, D.I. 1944. The Alturas floras. Carnegie Institution of Washington Publication, 553:263-284.
Bell, C.J., Lundelius Jr., E.L., Barnosky, A.D., Graham, R.W., Lindsay, E.H., Ruez Jr., D.R., Semken Jr., A., Webb, S.D., and Zakrzewski, R.J. 2004. The Blancan, Irvingtonian, and Rancholabrean Mammal Ages, p. 232-314. In Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic Mammals of North America. Columbia University Press, New York.
Borchardt, G.A. 1974. The SIMAN coefficient for similarity analysis. Classification Society Bulletin, 3:1-8.
Borchardt, G.A., Aruscavage, P.J., and Millard, H.T., Jr. 1972. Correlation of Bishop Ash, a Pleistocene marker bed, using instrumental neutron activation analysis. Journal of Sedimentary Petrology, 42:301-306.
Carmichael, E.S.E., Lange, R., Hall, C., and Renne, P. 2006. Faulted and tilted Pliocene olivine-tholeiite lavas near Alturas, NE California, and their bearing on the uplift of the Warner Range. Geological Society of America Bulletin, 118:1196-1211.
Collins, N.E. 1999. Stratigraphy and geochemistry of the Alturas Formation, Modoc Plateau, Northeastern California. Unpublished M.S. Thesis, Humboldt State University, CA.
Dorf, E. 1933. Pliocene floras of California. Carnegie Institution of Washington Publication , 412:1-112.
Evernden, J.E., Savage, D.E., Curtis, G.H., and James, G.T. 1964. Potassium-argon dates and the Cenozoic mammalian chronology of North America. American Journal of Science, 262:145-198.
Gradstein, F.M., Ogg, J.O., Schmitz, M., and Ogg, G. 2012. The Geologic Time Scale 2012. Elsevier B.V., Oxford, England.
Krebs, W.N., Bradburry, J.P., and Theriot, E. 1987. Neogene and Quaternary lacustrine diatom biochronology, western USA. Palaios, 2:505-513.
LaMotte, R.S. 1936. The Upper Cedarville flora of northwestern Nevada and adjacent California. Carnegie Institution of Washington Publication, 455:57-142.
Lindsay, E., Mou, Y., Downs, W., Pederson, J., Kelly, T., Henry, C., and Trexler, J. 2002. Recognition of the Hempillian/Blancan Boundary in Nevada. Journal of Vertebrate Paleontology, 22:429-442.
May, S.R., Woodburne, M.O., Lindsay, E.H., Albright, L.B., Sarna-Wojcicki, A., Wan, E., and Wahl, D.B. 2011. Geology and mammalian paleontology of the Horned Toad Hills, Mojave Desert, California. Palaeontologia Electronica, 14.3.28A:63pp, http://palaeo-electronica.org/2011_3/11_may/index.html
McKee, E.H., Duffield, W., and Stern, R. 1983. Late Miocene and early Pliocene basaltic rocks and their implications for crustal structure, northeastern California and south-central Oregon. Geological Society of America Bulletin, 94:292-304.
Morgan, L. and McIntosh, W. 2005. Timing and development of the Heise volcanic field, Snake River Plain Idaho, western USA. Geological Society of America Bulletin, 117:288-306.
Mou, Y. 1997. A new arvicoline species (Rodentia: Crietidae) from the Pliocene Panaca Formation, Southeast Nevada. Journal of Vertebrate Paleontology, 17:376-383.
Mou, Y. 1998. Schmelzmuster of Mimomys panacaensis. p. 79-90. In Tomida, Y.Y., Flynn, L.J., and Jacobs, L.L. (eds.), Advances in Vertebrate Paleontology and Geochronology. National Science Museum Monographs, No. 14, Tokyo, Japan.
Mou, Y. 2011. Cricetid rodents from the Pliocene Panaca Formation, southeastern Nevada, USA. Palaeontologia Electronica, 14.3.31A:53 pp, palaeo-electronica.org/2011_3/19_mou/index.html
Myers, A.T., Raymond, G.H., Connor, J.J., Conklin, N.M., and Rose, H.J., Jr. 1976. Glass reference standards for the trace-element analysis of geological materials - comparison of inter-laboratory data. U.S. Geological Survey Professional Paper, 1013:1-29.
Repenning, C.A. 1967. Paleartic-Nearctic mammalian dispersal in the late Cenozoic, p. 288-311. In Hopkins, D.M. (ed.), The Bering Land Bridge. Stanford University Press, Stanford, California.
Repenning, C.A. 1987. Biochronology of the microtine rodents of the United States, p. 236-268. In Woodburne, M.O. (ed.), Cenozoic Mammals of North America-Geochronology and Biostratigraphy. University of California Press, Berkeley, California.
Repenning, C.A. 2003. Chapter 17. Mimomys in North America. Bulletin American Museum of Natural History, 279:469-512.
Sarna-Wojcicki, A.M. 1971. Correlation of late Cenozoic pyroclastic deposits in the central Coast Ranges of California . Unpublished Ph. D. Dissertation, University of California, Berkeley.
Sarna-Wojcicki, A.M. 2000. Tephrochronology, p. 357-377. In Noller, J.S., Sowers, J.M., and Lettis, W.R. (eds.), Quaternary Geochronology: Methods and Applications. American Geophysical Union Reference Shelf 4.
Sarna-Wojcicki, A.M. and Davis, J.O. 1991. Quaternary Tephrochronology, p. 93-116. In Morrison, R.B. (ed.), Quaternary Nonglacial Geology: Conterminous United States. Geological Society of America, Decade of North American Geology Series, K-2, Boulder, Colorado.
Sarna-Wojcicki, A.M., Deino, A.L., Fleck, R.J., McLaughlin, R.J., Wagner, D., Wan, E., Wahl, D., Hillhouse, J.W., and Perkins, M. 2011. Age, composition, and areal distribution of the Pliocene Lawlor Tuff, and three younger Pliocene tuffs, California and Nevada. Geosphere, 7:599-628.
Sarna-Wojcicki, A.M., Bowman, H.R., Meyer, C.E., Russell, P.C., Woodward, M.J., McCoy, G., Rowe, J.J., Jr., Baedecker, P.A., Asaro, F., and Michael, H. 1984. Chemical analyses, correlations, and ages of upper Pliocene and Pleistocene ash layers of east-central and southern California. U.S. Geological Survey Professional Paper, 1293:40.
Sarna-Wojcicki, A.M., Reheis, M.C., Pringle, M.S., Fleck, R.J., Burbank, D., Meyer, C.E., Slate, J.L., Wan, E., Budahn, J.R., Troxel, B., and Walker, J.P. 2005. Tephra layers of Blind Spring Valley and related upper Pliocene and Pleistocene tephra layers, California, Nevada and Utah: Isotopic ages, correlation, and magnetostratigraphy. U.S. Geological Survey Professional Paper, 1701:63.
Wood, H.E. II, Chaney, R.W., Clark, J., Colbert, E.H., Jepsen, G.L., Reeside Jr., J.B., and Stock, C. 1941. Nomenclature and correlation of the North American continental Tertiary. Bulletin of the Geological Society of America, 52:1-48.
Woodburne, M.O. 1996. Precision and resolution in mammalian chronostratigraphy: Principles, practices, examples. Journal of Vertebrate Paleontology, 16:531-555.
Woodburne, M.O. 2004. Principles and Procedures, p. 1-20. In Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic Mammals of North America. Columbia University Press, New York.
