

 Unveiling trampling history through trackway interferences and track preservational features: a case study from the Bletterbach gorge (Redagno, Western Dolomites, Italy)
Unveiling trampling history through trackway interferences and track preservational features: a case study from the Bletterbach gorge (Redagno, Western Dolomites, Italy)
Article number: 19.2.20A
https://doi.org/10.26879/611
Copyright Palaeontological Association, July 2016
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 28 October 2015. Acceptance: 1 June 2016
{flike id=1499}
ABSTRACT
A slab from the Upper Permian Arenaria di Val Gardena Formation, bearing hundreds of tracks of different tetrapods, has been studied in order to reconstruct the trampling history through standard ichnological analysis and 3D photogrammetry. Analysis of crossed trackways to determine the relative timing of passages, as well as preservational features of the tracks qualitatively considered with trackmaker dimensions, has elucidated the nature of the trampled layer and the preservational mode of the tracks, especially the different conditions (i.e., sediment cohesion) that characterized the trampled layer during trackmaker crossings. Moreover, the analysis of an event timeline strongly suggests that caution should be exercised in analyzing track interferences, especially when multiple lines of evidence are lacking. As a consequence, the timeline of events may be masked, misleading and misinterpreted with obvious repercussions on general reconstructions (e.g., palaeoecological and behavioural). This study highlights the reliability of this type of analysis in untangling the preservational history of trampled surfaces; a useful investigative tool to be employed prior to traditional ichnological analyses aimed at classification of tetrapod tracks.
Paolo Citton. Dipartimento di Scienze della Terra, “Sapienza” Università di Roma, Piazzale Aldo Moro 5, 00185, Rome, Italy. paolo.citton@uniroma1.it
Iacopo Nicolosi. Istituto Nazionale di Geofisica e Vulcanologia, Via di Vigna Murata 605, 00143 Roma, Italy. iacopo.nicolosi@ingv.it
Roberto Carluccio. Istituto Nazionale di Geofisica e Vulcanologia, Via di Vigna Murata 605, 00143 Roma, Italy. roberto.carluccio@ingv.it
Umberto Nicosia. Dipartimento di Scienze della Terra, “Sapienza” Università di Roma, Piazzale Aldo Moro 5, 00185, Rome, Italy. umberto.nicosia@uniroma1.it
Keywords: Arenaria di Val Gardena; trackways; trackmakers; substrate; Late Permian
Final citation: Citton, Paolo, Nicolosi, Iacopo, Carluccio, Roberto, and Nicosia, Umberto. 2016. Unveiling trampling history through trackway interferences and track preservational features: a case study from the Bletterbach gorge (Redagno, Western Dolomites, Italy). Palaeontologia Electronica 19.2.20A: 1-20. https://doi.org/10.26879/611
palaeo-electronica.org/content/2016/1499-bletterbach-gorge-trackways
INTRODUCTION
The historical and idiographic nature of tetrapod ichnology, focusing on unique and unrepeatable events, is evident in the context of the dynamics of track formation. This process does not always occur in the same conditions, often leading to very different footprints largely depending on substrate properties as well as trackmaker anatomy and behaviour. As stated by several authors, the footprint itself should be considered as the result of in continuum interactions between the substrate and the trackmaker foot (e.g., Gatesy et al., 1999; Fornós et al., 2002; Gatesy, 2003; Milàn et al., 2006). The parts of the footprint and their formation can be considered “diachronous” in the time interval in which they were impressed (Romano et al., 2015). The physical properties (e.g., substrate consistency) of the tracking surface and the substrate exert a strong control on the footprint’s three-dimensional morphology, as highlighted by several papers that stress the specificity of some track features in relation to the moisture content of the substrate (e.g., Avanzini, 1998; Nadon, 2001; Milàn, 2006; Milàn and Bromley, 2006, 2008; Jackson et al., 2009, 2010; Bates et al., 2013).
Both of the main variables involved in the process of track formation have a prominent historical nature, but at very different timescales. The historical nature of the trackmaker and track is evident in the context of the irreversible process of evolution, considering the acquisition of anatomical and behavioural characters that are mirrored in the footprint’s three-dimensional morphology (Avanzini et al., 2001; Carrano and Wilson, 2001; Romano et al., 2015). The historical nature of the substrate, acting on a smaller timescale, involves variations in the physical and mechanical properties that can alternate or coexist on a trampled surface; these variations are detectable from footprint structure and substrate-related extramorphologies (for the concept of extramorphologies in tetrapod ichnology see Peabody, 1948; Haubold, 1996, 1998), as well as from other abiotic structures (e.g., Citton et al., 2012; Cariou et al., 2014). Thus, the identification and understanding of the main events and substrate conditions is deemed essential in order to reconstruct the dynamics of track formation.
Ancient surfaces with multiple tracks and trackways are quite common in the ichnological record; in these cases, the spatial distribution of tracks and trackways, as well as the trackmaker’s advancement direction, bending, interferences (i.e., interruptions and intersections) and overlapping, represent valuable data for ichnological analyses giving insights for general interpretations. A popular example is the recognition of herd behaviour in different groups of dinosaurs based on the analysis of parallel trackways (e.g., Currie and Sarjeant, 1979; Currie, 1983; Carpenter, 1992; Casanovas Cladellas et al., 1993; Barnes and Lockley, 1994; Cotton et al., 1998; Castanera et al., 2011), as well as the interpretation of interrupted trackways as evidence of predatory interactions between both vertebrate and invertebrate trackmakers in the continental Permian ichnological record (Lockley and Madsen, 1993; Hunt et al., 1994; Kramer et al., 1995, but see Hunt and Lucas, 1998 and Citton et al., 2012 for different interpretations).
At the same time, interferences between tracks and trackways have proved to be a significant source of direct and indirect information about trackmakers and substrate conditions during track formation (Lockley et al., 1986; Farlow et al., 1989; Mezga and Bajraktarevic, 1999; Matsukawa et al., 2001; Romero-Molina et al., 2003; Gangloff et al., 2004; Rainforth, 2005; Latorre Macarrón et al., 2006; Melchor and De Valais, 2006; Torcida Fernández-Baldor et al., 2006; Zhang et al., 2006; Nicosia et al., 2007; Wilson et al., 2009; Alcalá et al., 2012; Citton et al., 2012; Hornung et al., 2012; Razzolini et al., 2014), indirectly allowing reconstruction of the sequence of trackmaker passages.
Here, we analyze a single surface trampled by different types of tetrapods crossing in different directions. On the slab surface hundreds of tracks and traces are preserved. Ten trackways have been identified in addition to many isolated footprints and some probable invertebrate traces. Nine of the trackways and some Pachypes dolomiticus isolated tracks reciprocally interfere with one or more trackways by overprinting and/or intersections, allowing tracing back a relative timing of surface crossings. The analysis enabled a detailed narrative of events that is crucial for the understanding of the substrate conditions before, during and after the trampling, as well as for the recognition of the preservational mode of the tracks (Milàn and Bromley, 2006; Marty et al., 2009).
Geological Setting of the Arenaria di Val Gardena Formation in the Southern Alpine Domain
Two main, unconformity-bounded tectonosedimentary cycles have been recognized in the Late Carboniferous to early Middle Triassic stratigraphic succession of the Southern Alps (Massari et al., 1994). The lower cycle typically consists of volcanic rocks and alluvial to lacustrine sedimentary rocks deposited in small graben or half-graben. The upper cycle is characterized by an interruption of volcanic activity and is mainly represented by fluvial, coastal sabkha and shallow marine sediments (Cassinis et al., 1988). Radiometric data and palynomorph assemblages (Cassinis and Doubinger, 1992; Cassinis et al., 2002) indicate an Artinskian-Kungurian age for the lower cycle (Early Permian), whereas the upper cycle is regarded as Wuchiapingian (Late Permian) on the basis of sporomorphs and foraminifera (Cassinis et al., 2002).
The arenaceous and silty terrestrial red beds of the Arenaria di Val Gardena Formation, first described by von Richthofen (1860), belong to the second cycle and can lie directly on the crystalline basement, on conglomeratic lenses (Bosellini and Dal Cin, 1968) or on volcanic rocks of the “Gruppo Vulcanico Atesino” (Avanzini et al., 2008). The unit is laterally and upwardly replaced by the shallow marine Bellerophon Formation related to westward transgressions, which is in turn overlain by the Lower Triassic Werfen Formation (Cassinis et al., 2012; Roghi et al., 2014).
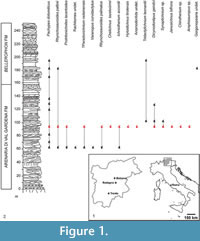 One of the most important localities to study the Late Permian evolution of the Southern Alpine domain is, undoubtedly, the Butterloch-Bletterbach section, close to the town of Redagno (Bolzano, Italy; Figure 1.1), where a thick sequence of the Arenaria di Val Gardena Formation and Bellerophon Formation crops out. The Bletterbach gorge was extensively studied in the past and more recently, with regard to sedimentological and stratigraphical features (e.g., Accordi, 1958; Conti et al., 1986, 1997; Massari et al., 1988; Cassinis et al., 1999, 2002, 2012), the palaeobotanical and palynological associations (e.g., Leonardi, 1948; Klaus, 1963; Kustatscher et al., 2012, 2014; Bauer et al., 2014) and the rich tetrapod footprint ichnofauna (e.g., Kittl, 1891; Abel, 1929; Leonardi, 1951; Leonardi and Nicosia, 1973; Leonardi et al., 1975; Conti et al., 1975, 1977, 1979; Valentini et al., 2007, 2008, 2009).
One of the most important localities to study the Late Permian evolution of the Southern Alpine domain is, undoubtedly, the Butterloch-Bletterbach section, close to the town of Redagno (Bolzano, Italy; Figure 1.1), where a thick sequence of the Arenaria di Val Gardena Formation and Bellerophon Formation crops out. The Bletterbach gorge was extensively studied in the past and more recently, with regard to sedimentological and stratigraphical features (e.g., Accordi, 1958; Conti et al., 1986, 1997; Massari et al., 1988; Cassinis et al., 1999, 2002, 2012), the palaeobotanical and palynological associations (e.g., Leonardi, 1948; Klaus, 1963; Kustatscher et al., 2012, 2014; Bauer et al., 2014) and the rich tetrapod footprint ichnofauna (e.g., Kittl, 1891; Abel, 1929; Leonardi, 1951; Leonardi and Nicosia, 1973; Leonardi et al., 1975; Conti et al., 1975, 1977, 1979; Valentini et al., 2007, 2008, 2009).
The rich track-bearing levels belong to a stratigraphic interval that shows a wide range of facies, as highlighted by Conti et al. (1986; Figure 1.2). According to Massari et al. (1988), the spectrum comprises channel-fill sandstones, red to grey mudstones, small thin-bedded intervals of sandstones, commonly wave-rippled at the top, and mudstones sometimes cracked, crinkled and with gypsum nodules and, finally, laminated and bioturbated siltstones and fine sandstones; intervals with pedogenic properties, such as carbonate nodules or concretions, have also been observed. The facies associations have been related to the transition between the distal portion of a fluvial system and a coastal sabkha that was susceptible to floods by very shallow water, as suggested by the thin-bedded sequence of wave-rippled fine sandstones with mud drapes (Conti et al., 1986; Massari et al., 1988, 1994).
MATERIAL AND METHODS
Much of the terminology used here was introduced and discussed by Leonardi (1987). The study of the relative timing of passages, main advancement directions and preservational features of the tracks were carried out directly on the slab. A grid, consisting of three rows and eight columns indicated with letters from A to C and Arabic numerals from 0 to 7, respectively, has been superimposed on the interpretive drawing to facilitate the description of the main intersection points and interferences between tracks. Each trackway is indicated by an acronym that is followed (only for trackways of the same ichnotaxon) by an Arabic numeral according to the relative time of formation. The acronyms are: ‘Gl1-7’ for Ganasauripus ladinus tracks and trackways, ‘Jb’ for Janusichnus bifrons trackway, ‘Ct1-2’ for Chelichnus tazelwürmi trackways and ‘Pd’ for Pachypes dolomiticus tracks and trackway. Sedimentary structures are indicated by the acronyms ‘rip’ (i.e., ripple marks) and ‘dcr’ (i.e., possible desiccation cracks).
The slab MPUR NS 34/28 (MPUR NS: Museo di Paleontologia Università di Roma Nuova Serie) was collected in the 1980s from the Butterloch-Bletterbach section (Ceoloni et al., 1988). Digital 3D models of MPUR NS 34/28 were constructed in order to obtain a quantitative assessment of the relief of the studied tracks (qualitatively evaluated during the firsthand analysis). The model was achieved using the high resolution Digital Photogrammetry method. This technique is based on Structure from Motion (SfM) (Ullman, 1979) and Multi View Stereo (MVS) (Seitz et al., 2006) algorithms and takes advantage the computational power of modern computers, low cost digital cameras and high quality photos.
A full 3D scaled model of any object in space can be reconstructed, starting from a suitable number of photographic images (2D object projection) of a static scene taken from different points of view. To achieve suitable results for scientific applications, it is crucial that objects possess locally unique texture, and numerous photos are taken in order to maintain a high “overlay redundancy index” and a small angular separation between images. The former index can be thought as the average number of photos in which surveyed surface points are present. This technique produces high-quality, dense point clouds with no spatial information that can be used to generate 3D models comparable to those obtained from LiDAR technology.
A number of software solutions do exist for digital photogrammetry; the software package used (Agisoft PhotoScan Pro, www.agisoft.com) is geared toward semi-automatic processing of images with the end goal of the creation of 3D textured meshes. PhotoScan can handle a wide range of digital images and automatically calculates camera calibrations.
The photogrammetric process used images that were acquired using an 8 Megapixel Canon Digital ixus 80 IS and a 10 Megapixel Canon EOS 400D with 18 mm focal length. In order to achieve good image overlap, 270 images were used in 3D model construction. In order to correctly scale the calculated model, a metric reference marker was applied on the surface. A dense cloud mean resolution of less than 1 mm was obtained.
RESULTS
The Slab
 The slab is composed of about 20 parts, some of which were obtained by gluing together blocks of small sizes. When reconstructed, the slab is about 2.5 m long, a maximum of 1 m wide and about 3 cm thick (Figure 2). The slab is composed of a dark-red siltstone. Tracks and traces are preserved as convex hyporeliefs, thus representing the infills of the original impressions. In section, well below the alleged upper limit of the filling depositional event, silty sediments are characterized by very fine, undeformed, parallel laminae. Sediments constituting the filling of the tracks are instead unstructured, suggesting that the burial of the tracking surface (sensu Fornós et al., 2002) likely occurred in a relatively rapid sedimentation event forming natural casts (see “track burial model” in Marty et al., 2009). The surface on which the tracks are preserved is slightly undulatory and characterized by weakly developed, discontinuous, straight-crested ripple marks. The ripple marks are symmetrical and were likely produced by oscillatory flow acting, after a flooding event, on silty sediments. The discontinuity of the ripples can be ascribed to an original topographical heterogeneity of the tracked surface.
The slab is composed of about 20 parts, some of which were obtained by gluing together blocks of small sizes. When reconstructed, the slab is about 2.5 m long, a maximum of 1 m wide and about 3 cm thick (Figure 2). The slab is composed of a dark-red siltstone. Tracks and traces are preserved as convex hyporeliefs, thus representing the infills of the original impressions. In section, well below the alleged upper limit of the filling depositional event, silty sediments are characterized by very fine, undeformed, parallel laminae. Sediments constituting the filling of the tracks are instead unstructured, suggesting that the burial of the tracking surface (sensu Fornós et al., 2002) likely occurred in a relatively rapid sedimentation event forming natural casts (see “track burial model” in Marty et al., 2009). The surface on which the tracks are preserved is slightly undulatory and characterized by weakly developed, discontinuous, straight-crested ripple marks. The ripple marks are symmetrical and were likely produced by oscillatory flow acting, after a flooding event, on silty sediments. The discontinuity of the ripples can be ascribed to an original topographical heterogeneity of the tracked surface.
Brief Description of Ichnotaxa
The studied slab has been previously described with regard to ichnotaxonomy and trackmaker identification (Ceoloni et al., 1988; Valentini et al., 2007, 2008, 2009). A general description of the studied tracks is briefly provided; for a detailed discussion of ichnotaxonomy, which is beyond the aim of this paper, reference should be made to the cited literature and references therein.
 According to Ceoloni et al. (1988), Chelichnus tazelwürmi refers to footprints with a marked affinity with ‘mammalian reptiles’ belonging to ‘pelycosaurs’ (Figure 3.1-2). Manus and pes are pentadactyl, semiplantigrade and mesaxonic, the pes with a marked internal functional prevalence; digit traces are straight and almost parallel to each other and relative to the trackway midline (Ceoloni et al., 1988).
According to Ceoloni et al. (1988), Chelichnus tazelwürmi refers to footprints with a marked affinity with ‘mammalian reptiles’ belonging to ‘pelycosaurs’ (Figure 3.1-2). Manus and pes are pentadactyl, semiplantigrade and mesaxonic, the pes with a marked internal functional prevalence; digit traces are straight and almost parallel to each other and relative to the trackway midline (Ceoloni et al., 1988).
Ceoloni et al. (1988) also established the ichnotaxon Janusichnus bifrons (Figure 3.3) for pentadactyl, plantigrade manus and pes footprints. The pes shows some ‘mammalian’ features, among which a reduced phalangeal formula and internal functional prevalence are the most prominent. The manus has digits increasing in length from I to IV, with the fifth reduced (Ceoloni et al., 1988).
Ganasauripus ladinus was established by Valentini et al. (2007) as part of the review of the lacertoid footprints of the Arenaria di Val Gardena Formation. It indicates pentadactyl, semiplantigrade to plantigrade footprints of a quadruped ‘reptile’, showing a mesaxonic manus and an ectaxonic pes and sinusoidal tail drag marks; digits increase in length from I to IV, with the fifth ending behind the base of digit IV (Figure 3.4, Figure 4.1).
 Pachypes dolomiticus was coined by Leonardi et al. (1975) and recently re-evaluated by Valentini et al. (2009). The ichnotaxon refers to semiplantigrade, pentadactyl footprints of a quadruped trackmaker identified as a pareiasaur (e.g., Valentini et al., 2008, 2009). The pes, sub-parallel with respect to the trackway midline, exhibits a total functional prevalence, whereas the manus is mesaxonic and often turned inward with respect to the trackway midline. Digits are short and sturdy, sub-parallel; external digits are shorter (Figure 4.2-4).
Pachypes dolomiticus was coined by Leonardi et al. (1975) and recently re-evaluated by Valentini et al. (2009). The ichnotaxon refers to semiplantigrade, pentadactyl footprints of a quadruped trackmaker identified as a pareiasaur (e.g., Valentini et al., 2008, 2009). The pes, sub-parallel with respect to the trackway midline, exhibits a total functional prevalence, whereas the manus is mesaxonic and often turned inward with respect to the trackway midline. Digits are short and sturdy, sub-parallel; external digits are shorter (Figure 4.2-4).
Event Timeline
The analysis of interferences between trackways, and between tracks and sedimentary structures, allows us to define a relative timeline for when different crossing events occurred (Figure 5.1-9). Gl3 tracks and some isolated tracks do not interfere with any other trace, so their relative timing by means of intersection points cannot be defined. Footprints Pdx and Pdy (Figure 5.8-9) were not considered in the analysis due to their poor quality and the uncertainty in referring them confidently to any single trackway.
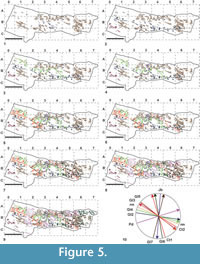 The formation of ripple marks represents the most likely first event recorded on the submerged surface, as indicated by their interference by some tetrapod tracks (Figure 5.1, meshes B5-B6; e.g., Figure 6). In many cases, ripple crests are interrupted at track margins or, when present, at the lateral edge of the expulsion rims.
The formation of ripple marks represents the most likely first event recorded on the submerged surface, as indicated by their interference by some tetrapod tracks (Figure 5.1, meshes B5-B6; e.g., Figure 6). In many cases, ripple crests are interrupted at track margins or, when present, at the lateral edge of the expulsion rims.
 According to Valentini et al. (2007), Gl1 tracks, assigned to Ganasauripus ladinus (meshes B3-B4, Figure 5.1), exhibit a more digitigrade posture of the trackmaker’s feet and lack impressions of the first and the fifth digits; digit traces are widened and have claw scratch marks (Figure 7). These features characterize Gl1 tracks as swimming traces (Valentini et al., 2007). According to the same authors, these tracks were likely produced by a trackmaker that was moving on a thin sheet of water partially floating, just touching the substrate surface by pushing off with the distal ends of digit.
According to Valentini et al. (2007), Gl1 tracks, assigned to Ganasauripus ladinus (meshes B3-B4, Figure 5.1), exhibit a more digitigrade posture of the trackmaker’s feet and lack impressions of the first and the fifth digits; digit traces are widened and have claw scratch marks (Figure 7). These features characterize Gl1 tracks as swimming traces (Valentini et al., 2007). According to the same authors, these tracks were likely produced by a trackmaker that was moving on a thin sheet of water partially floating, just touching the substrate surface by pushing off with the distal ends of digit.
 After the Gl1 trackmaker, the animal that produced trackway Gl2, and likely the Gl3 tracks, crossed the surface (Figure 5.2, meshes B0-B6). The tetrapod, which produced 13 strides and a faint tail trace, interfered with some ripple marks (mesh C6). The relative antecedence of this crossing is derived by different points of intersection between the trackway Gl2 and the trackways Gl4, Gl5, Gl6, Gl7 and Pd (see Figure 5.9, meshes B0-B2 and C3-C6, and Figure 5.10).
After the Gl1 trackmaker, the animal that produced trackway Gl2, and likely the Gl3 tracks, crossed the surface (Figure 5.2, meshes B0-B6). The tetrapod, which produced 13 strides and a faint tail trace, interfered with some ripple marks (mesh C6). The relative antecedence of this crossing is derived by different points of intersection between the trackway Gl2 and the trackways Gl4, Gl5, Gl6, Gl7 and Pd (see Figure 5.9, meshes B0-B2 and C3-C6, and Figure 5.10).
Later, the producer of the Gl4 trackway passed (Figure 5.3, meshes A1-C5). The interferences preserved on the slab indicate the relative antecedence of this trackway to the Gl5, Gl6, Jb and Pd trackways (Figure 5.9-10, meshes B2-B4; Figure 8.1). Subsequent to Gl4, the Jb trackmaker crossed the surface (Figure 5.4, meshes A3-B3), trampling on the tail trace and a pes print of the Gl4 trackway. The Jb trackway disappears at the intersection with a pes impression belonging to the Pd trackway, thus indicating its relative subsequence.
Before the impression of the Gl5 trackway, the trackmaker of Ct1 left about 10 poorly preserved footprints (Figure 5.5, mesh A1-B1). The Gl5 trackway, made of about 14 sets and by a sinusoidal tail trace, was left by a trackmaker (Figure 5.5, meshes A0-C3) interfering with the Gl2, Gl4 and Ct1 trackways (Figure 5.9-10,  mesh A1-B1). Trackway Ct2 was produced before Gl6 and Gl7. While crossing the surface, the Ct2 trackmaker interfered with ripple marks, some swimming traces and with the trackway Gl2 (Figure 5.6, meshes B4-B5 and C5-C6). Afterward, the Gl6 trackmaker crossed the surface, interfering with the Gl2, Gl4 and Ct2 trackways (Figure 5.7, mesh A5-C5).
mesh A1-B1). Trackway Ct2 was produced before Gl6 and Gl7. While crossing the surface, the Ct2 trackmaker interfered with ripple marks, some swimming traces and with the trackway Gl2 (Figure 5.6, meshes B4-B5 and C5-C6). Afterward, the Gl6 trackmaker crossed the surface, interfering with the Gl2, Gl4 and Ct2 trackways (Figure 5.7, mesh A5-C5).
Trackway Gl7 is comprised of six sets, two of which are incomplete, and a sinusoidal tail trace. The penultimate pes impression of this trackway shows a trace probably left by the tarsal-ankle joint pad, as highlighted by Valentini et al. (2007). Regarding the relative timeline, it can only be stated that this trackway was left by a trackmaker crossing the surface after the impression of the Gl2 and Ct2 trackways, with which it interferes (Figure 5.7, mesh B5), and before the Pd trackway.
The last trackmaker leaving traces of its passage was the producer of the Pd trackway, inferred to be a member of Pareiasauridae (Valentini et al., 2008; Avanzini et al., 2011). The trackway is made of three sets, the first of which is incomplete. The trackmaker crossed the surface and interfered with ripples marks and the Gl2, Gl4, Gl5, Gl6, Gl7 and Jb trackways (Figure 5.8-10, meshes A6-B1; Figure 8.2).
The tracked surface also possesses structures tentatively interpreted as desiccation cracks (indicated as dcr in Figure 5.9-10, meshes B5, B6), very likely formed well after track formation.
Preservational Features
The tracks are not uniformly preserved on the surface; instead preservational variations can be observed between and within trackways. At the same time, many of the natural casts clearly show some recurrent features providing significant information about the substrate conditions during and after trampling. All the tracks seem to have been impressed at the same depth, regardless of footprint morphology and trackmaker anatomy. Maximum variation in track relief is within a centimetre. Tracks with greatest relief are those left by the pareiasaurian, as illustrated in the 3D photogrammetric model (Figure 9.2, Appendix).
 Trackways Gl4, Gl5, Gl6, Ct2, Pd and, with lesser emphasis, Gl7 and Jb, are generally characterized by very well-preserved footprints showing several anatomical characters of the trackmakers’ foot, as also stated by Valentini et al. (2007) for Ganasauripus ladinus trackways. In many cases footprints belonging to these trackways exhibit expulsion rims extending up to the proximal margin of the footprint. The rims are almost vertical, high and sharp. Track walls (sensu Brown, 1999) and lateral walls of digits are vertical, and no traces of collapse are observed. These features suggest that trampled sediments were highly cohesive and plastic during trackmaker crossings. In contrast, Gl1 footprints are characterized by weak expulsion rims and are more developed horizontally, suggesting that sediments were less cohesive and more fluid, likely due to water logging, during trackmaker advancement.
Trackways Gl4, Gl5, Gl6, Ct2, Pd and, with lesser emphasis, Gl7 and Jb, are generally characterized by very well-preserved footprints showing several anatomical characters of the trackmakers’ foot, as also stated by Valentini et al. (2007) for Ganasauripus ladinus trackways. In many cases footprints belonging to these trackways exhibit expulsion rims extending up to the proximal margin of the footprint. The rims are almost vertical, high and sharp. Track walls (sensu Brown, 1999) and lateral walls of digits are vertical, and no traces of collapse are observed. These features suggest that trampled sediments were highly cohesive and plastic during trackmaker crossings. In contrast, Gl1 footprints are characterized by weak expulsion rims and are more developed horizontally, suggesting that sediments were less cohesive and more fluid, likely due to water logging, during trackmaker advancement.
A different substrate condition can be inferred from the crossing of the trackmakers of Gl2 and Gl3. These trackways share similar preservational features; the right side of both trackways is not preserved, whereas the left one is incomplete, being made of shallowly impressed sets and often lacking the manus impressions. In these cases, the preservational bias causing the failure of the impression of the right side was likely controlled by different substrate conditions, in a restricted space.
Trackway Ct2 shows the greatest internal preservational variability. In the first segment of the trackway, like trackway Ct1, footprints are sometimes faintly preserved and sets are incomplete, although some retain the main anatomical characters. From the fifteenth set up to the end of the trackway, however, footprints are very well preserved and are characterized by almost vertical walls and definite expulsion rims. These features again suggest a wet and cohesive substrate during trackmaker crossing, at least for the central and final portion of the trackway.
A noteworthy feature is represented by the interferences that trackway Pd forms with some ripple marks and with Gl7, further indicating the conditions of the surface during the crossing of the huge trackmaker. In four cases, ripple marks overlap different Pachypes dolomiticus footprints (e.g. Figure 4.3-4; see Figure 5.8, meshes C2, C3; B5, B6), thus seemingly subsequent in time.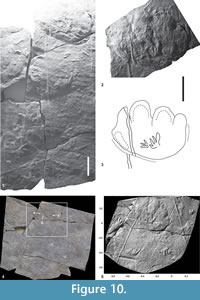 However, the antecedence of ripple marks is assured by the slight displacement affecting these structures at the intersections with the Pd tracks (e.g., mesh B5, B6 in Figure 5.8). Therefore, it is evident that the heavy trackmaker crossed the surface after the ripples formed, causing dislocation and overprinting, without obliterating the trampled structures.
However, the antecedence of ripple marks is assured by the slight displacement affecting these structures at the intersections with the Pd tracks (e.g., mesh B5, B6 in Figure 5.8). Therefore, it is evident that the heavy trackmaker crossed the surface after the ripples formed, causing dislocation and overprinting, without obliterating the trampled structures.
A similar configuration is observable between the Pd and Gl7 trackways (Figure 10.1). The tail trace of Gl7 is clearly interrupted in correspondence to the proximal margin of the foreprint of set 6 of Pd, but it can still be observed as superimposed on the Pd manus with a right displacement of less than 1 cm. The last set of Gl7 is preserved close to the tail trace and at the same distance as in the preceding sets, although it appears smaller than the previous sets (Figure 10.2-5). As in the case of ripple marks, the Pd trackmaker crossed the surface after the passage of the Gl7 trackmaker. As the giant animal walked and interfered with the Gl7 trackway, the tail trace was interrupted and dislocated, but at the same time the overprinted structures were preserved under the animal’s weight. Similar configuration is shown by the preceding pes impression of Gl7 (see Valentini et al., 2007 for a discussion of this track) and the pes impression of the Pd trackway (Figure 5.8, mesh A5, B5).
DISCUSSION
The conducted analyses enabled important information about footprint preservation and substrate conditions during and after the tetrapods’ crossings to be gained. By integrating the collected evidence in a broader perspective, additional interpretations can be made.
The configuration of Pd interferences with ripple marks and Gl7 trackways (i.e., interruption and displacement plus lowering and overprinting) requires an explanation that accounts for both the observed track preservational features and substrate properties during track formation. The observed interferences can occur within the substrate, with the trackmaker of Pd walking well above the trampled surface (i.e., Pd tracks should be the filling of undertracks or transmitted tracks or ghost prints sensu Marty et al., 2009). However, the lack of a stack of transmitted prints and no sediment interposition (nor differential compaction) between the fillings of Ganasauripusladinus and Pachypesdolomiticus suggests that there were no important sedimentation events between the two crossings. Thus Pd tracks are likely not undertracks, but natural casts similar to the other tracks on the surface. The most parsimonious explanation for Pd-Gl7 interference is that when the Pd trackmaker crossed the surface, the trampled sediments were deformed under the trackmaker’s weight and, simultaneously, the ripple crests, Gl7 tracks and the tail trace were lowered and displaced without being distorted, fractured or obliterated. At the time of the impression of the Pd trackway, some tracks were still so fresh as to be effaced (e.g., Jb), whereas the sediments affected by ripple marks and already trampled by other tetrapods probably had expelled part of the water. The surface hardened, allowing for the preservation of some ripple marks, the tail trace and the last set of the Gl7 trackway during the Pd trackmaker passage. The surface was still relatively plastic and ductile enough, however, not to collapse or crack under the pressures applied by the heavy trackmaker whose tracks are rather detailed. It can be envisaged that differential hardening of the substrate may have been triggered by progressive general dehydration of the sediments or by microbial activity. At the same time, small time scale variations of the sediment moisture, which may have been driven by the daily variation in humidity or rainfall, could also have occurred, thus explaining the co-existence of slightly different substrate conditions during Pd trackmaker crossing.
Moreover, the facts that i) tracks left by trackmakers of different relative sizes, morphologies and modes of locomotion, which crossed the same surface under different substrate conditions, were impressed to the same depth and that ii) track elements and substrate-related extramorphologies (e.g., anatomical details, track walls, expulsion rims, associated traces) of these tracks co-occur and share similar features, suggest that i) a subsurface consolidated layer underlay the trampled sediments, as also suggested by Valentini et al. (2007, see also Falkingham et al., 2011), and that ii) all the tracks were likely produced within days or, perhaps, weeks of each other. If trackmakers of different weights crossed the surface simultaneously under homogeneous substrate conditions, then the footprints produced would probably have different characteristics in relation to different pressure-sediment relationships, regardless of the existence of a firm subsurface layer.
A general consideration regards the overprinting of tracks and the clear difficulties in establishing the correct timeline of passages, especially when subsidiary evidences (i.e., more than one intersection point, interruption, displacement) are lacking or not observed. A vivid example is the overprinting between Gl7 and Pd tracks discussed above. In this case, assuming no record of interruption and displacement (e.g., trackmaker’s tail lifted from the ground during locomotion; substrate not compliant to record displacement), the reverse timing between the two crossings would probably have been interpreted as correct.
Finally, the maximum sediment thickness able to compact during the trampling history was roughly equal to that showed by the overprinted cases (e.g., the sum of Pd manus plus Gl7 set thickness), thus shedding further light on the role of fossil tracks as paleo-penetrometers (see Falkingham et al., 2011). Pd being the last set of tracks preserved along with the displaced tail trace of Gl6, it also possible to infer that during its trampling, the pareiasaurian trackmaker likely never reached the alleged consolidated layer underlying the trampled sediments.
CONCLUSIONS
The analysis carried out on a rich Late Permian ichnocoenosis preserved on the studied slab indicates a complex trampling history involving at least four different types of trackmaker crossing the surface during different times, in different directions, and under variable conditions of substrate consolidation. Interferences between tracks and trackways, as well as several track features and some substrate-related extramorphologies, have proved to be a meaningful reservoir of information about the timeline of events and the track preservational history.
The tracks, preserved as convex hyporeliefs, are natural casts representing the filling of footprints left by tetrapods on the tracking surface. Tracks were left over days or weeks, during which time the surface passed through different conditions with respect to sediment cohesion and hardening. At the same time, minor differences in substrate conditions probably occurred simultaneously and in a restricted space, ruling the dynamics of track formation and the variability within single trackways. Optimal conditions for track recording, with respect to trackmakers’ weight, occurred at different times during the trampling history. At the time of the crossing of the Pd trackmaker, the substrate was still in optimal condition with respect to the trackmaker’s weight to record the passage of the heavier producer, but likely not the lighter ones. The progressive hardening of the trampled layer, still without achieving desiccation, enabled the preservation of those tracks and traces, which were lowered and displaced under the limbs of the giant.
The analysis of interferences has stressed the importance of secondary evidence when establishing the relative antecedence or subsequence of events. Caution should be exercised in general reconstructions based on track interferences alone. In the studied case, the tracks of the pareiasaurian trackmaker turned out to be crucial in refining the timeline of events and in further constraining the trampling history as well as the original thickness of the trampled layer.
The need to deeply understand, case-by-case, the close interdependence between the momentary substrate conditions, trackmaker morphology, dimensions and behaviour, and the 3D track morphology is highlighted. This understanding is pivotal in the context of tetrapod ichnology, as it allows the direct investigation of the processes leading to track formation. In this regard, this type of analysis is strongly recommended whenever possible. While time-intensive, it could be a useful investigative tool prior to traditional ichnological analyses, for which the control on the mode of preservation of the footprints is crucial.
ACKNOWLEDGEMENTS
M. Romano is kindly thanked for comments on an early version of the manuscript. C. Leoncini and F. Battista are thanked for their help during museum operations. The Executive Editor (J. Louys), the Handling Editor (D. Hembree), the Style Editor (C. Grohe) and the reviewers are acknowledged for their comments and suggestions that greatly improved the manuscript.
REFERENCES
Abel, O. 1929. Eine Reptilien Fährte aus dem Oberen Perm (Grodener Sandstein) der Gegend von Bozen in Sudtirol. Paleobiology, 2:1-14.
Accordi, B. 1958. Contributo alla conoscenza del Permiano medio-superiore della zona di Redagno (Bolzano). Annali dell’Università di Ferrara, Nuova Serie, 3:37‐47.
Alcalá, L., Cobos, A., Espílez, E., Gascó, F., Mampel, L., Escorza, C.M., and Royo-Torres, R. and 2012. Ichnitas de dinosaurios en la Formación Villar de Arzobispo de Ababuj (Teruel, España). Geogaceta, 51:35-38.
Avanzini, M. 1998. Anatomy of a footprint: bioturbation as a key to understanding dinosaur walk dynamics. Ichnos, 6:129-139.
Avanzini, M., Bernardi, M., and Nicosia, U. 2011. The Permo-Triassic tetrapod faunal diversity in the Italian Southern Alps, p. 591-608. In Dar, I.A. and Dar, M.A. (eds.), Earth and Environmental Sciences. InTech, Rijeka.
Avanzini, M., Ceoloni, P., Conti, M.A., Leonardi, G., Manni, R., Mariotti, N., Mietto, P., Nicosia, U., Santi, G., and Spezzamonte, M. 2001. Permian and Triassic tetrapod ichnofaunal units of N. Italy, their potential contribution to continental biochronology, p. 89-107. In Cassinis, G. (ed.), Permian and continental deposits of Europe and other areas. Regional reports and correlations. Natura Bresciana, Annali del Museo Civico di Scienze Naturali di Brescia, Monografia 25.
Avanzini, M., Neri, C., Nicosia, U., and Conti, M.A. 2008. A new Early Permian ichnocoenosis from the “Gruppo vulcanico atesino” (Mt. Luco, Southern Alps, Italy). Studi Trentini di ScienzeNaturali Acta Geologica, 83:231-236.
Barnes, F.A. and Lockley, M.G. 1994. Trackway evidence for social sauropods from the Morrison Formation, Eastern Utah (USA). Gaia, 10:37-41.
Bates, K.T., Savage, R., Pataky, T.C., Morse, S.A., Webster, E., Falkingham, P.L., Ren, L., Qian, Z., Collins, D., Bennet, M.R., McClymont, J., and Crompton, R.H. 2013. Does footprint depth correlate with foot motion and pressure? Journal of the Royal Society Interface 10, 20130009.
Bauer, K., Kustatscher, E., Butzmann, R., Fischer, T.C., van Konijnenburg-van Cittert, J.H.A., and Krings, M. 2014. Gynkgophytes from the Upper Permian of the Bletterbach Gorge. RivistaItaliana di Paleontologia e Stratigrafia, 120:271-279.
Bosellini, A. and Dal Cin, R. 1968. Sedimentologia delle Arenarie di Val Gardena - Sezione di S. Martino in Badia (Bolzano). Bollettino della Società Geologica Italiana, 87:401-423.
Brown, T. Jr. 1999 . The science and art of tracking-nature’s path to spiritual discovery. Berkley Books, New York.
Cariou, E., Olivier, N., Pittet, B., Mazin, J.-M., and Hantzpergue, P. 2014. Dinosaur track record on a shallow carbonate-dominated ramp (Loulle section, Late Jurassic, French Jura). Facies, 60:229-253.
Carpenter, K. 1992. Behavior of hadrosaurs as interpreted from footprints in the “Mesaverde” Group (Campanian) of Colorado, Utah, and Wyoming. Contributions to Geology, 29:81-96.
Carrano, M.T. and Wilson, J.A. 2001. Taxon distributions and the tetrapod track record. Paleobiology, 27:564-582.
Casanovas Cladellas, M.L., Ezquerra Miguel, R., Fernandez Ortega, A., Perez-Lorente, F., Santafe Llopis, J.V., and Torcida Fernanadez, F. 1993. Track of a herd of webbed ornithopods and other footprints found in the same site (Igea, La Rioja. Spain). Revue de Paléobiologie, Special Volume 7:29-36.
Cassinis, G., Cortesogno, L., Gaggero, L., Massari, F., Neri, C., Nicosia, U., and Pittau, P (eds). 1999. Stratigraphy and facies of the Permian deposits between Eastern Lombardy and the Western Dolomites. Field Trip Guidebook . International Field Conference on “The Continental Permian of the Southern Alps and Sardinia (Italy). Regional Reports and General Correlations”. Earth Science Department, Pavia University, Italy.
Cassinis, G. and Doubinger, J. 1992. Artinskian to Ufimian palynomorph assemblages from the Central Southern Alps (Italy) and their regional stratigraphic implications. In Nairn, A.E.M. and Koroteev, V. (eds.), Contributions to Eurasian Geology. Occasional Publication ESRI New Series 8b, Part I. Columbia University of South Carolina, South Carolina, USA.
Cassinis, G., Massari, F., Neri, C., and Venturini, C. 1988. The continental Permian in the Southern Alps (Italy). A review. Zeitschrift für Geologische Wissenschaften, 16:1117-1126.
Cassinis, G., Nicosia, U., Pittau, P. and Ronchi, A. 2002. Palaeontological and radiometric data from the Permian continental deposits of the central-eastern southern Alps (Italy), and their stratigraphic implications. Mémoire de l’Association des Géologues du Permien, 2:53-74.
Castanera, D., Barco, J.L., Díaz-Martínez, I., Herrero Gascón, J., Pérez-Lorente, F., and Canudo, J.I. 2011. New evidence of a herd of titanosauriform sauropods from the lower Berriasian of the Iberian range (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 310:227-237.
Ceoloni, P., Conti, M.A., Mariotti, N., and Nicosia, U. 1988. New Late Permian tetrapod footprints from Southern Alps. Memorie della Società Geologica Italiana, 34:45-65.
Citton, P., Sacchi, E., and Nicosia, U. 2012. Sometimes they come back: recovery and reinterpretation of a trackway slab from the Permian Coconino Sandstone of the Southwestern United States. Ichnos, 19:165-174.
Conti, M.A., Fontana, D., Mariotti, N., Massari, F., Neri, C., Nicosia, U., Pasini, M., and Pittau, D. 1986. The Bletterbach-Butterloch section (Val Gardena Sandstone and Bellerophon Formation), p. 99-119. In Italian IGCP 203 Group (eds.), Field conference on Permian and Permian-Triassic boundary in the South-Alpine segment of the Western Tethys. Field Guide-Book. Società Geologica Italiana, Brescia, Italy .
Conti, M.A., Leonardi, G., Mariotti, N., and Nicosia, U. 1975. Tetrapod footprints, fishes and molluscs from the Middle Permian of the Dolomites (N. Italy). Memorie Geopaleontologiche,Università di Ferrara, 3:141-150.
Conti, M.A., Leonardi, G., Mariotti, N., and Nicosia, U. 1977. Tetrapod footprints of the “Arenaria di Val Gardena” (North Italy). Their paleontological, stratigraphic and palaeoenvironmental meaning. Paleontographia Italica, LXX (serie XL):1-91.
Conti, M.A., Leonardi, G., Mariotti, N., and Nicosia, U. 1979. A review of a group of Permian tetrapod footprints from Northern Italy. Memorie di Scienze Geologiche, XXXIV:143-152.
Conti, M.A., Mariotti, N., Nicosia, U., and Pittau, P. 1997. Succession of selected bioevents in the continental Permian of the Southern Alps (Italy): improvements of intrabasinal and interregional correlations, p. 51-65. In Dickins, J.M., Yang, Z.Y., Yin, H.F., Lucas, S.G., and Acharya, S.K. (eds.), Late Palaeozoic and Early Mesozoic Circum-Pacific Events and Their Global Correlation. Cambridge University Press, World and Regional Geology Series 10.
Cotton, W., Cotton, J., and Hunt, A. 1998. Evidence for social behavior in ornithopod dinosaurs from the Dakota Group of northeastern New Messico, U.S.A. Ichnos, 6:141-149.
Currie, P.J. 1983. Hadrosaur trackways from the Lower Cretaceous of Canada. ActaPalaeontologica Polonica, 28:63-73.
Currie, P.J. and Sarjeant, W.A.S. 1979. Lower Cretaceous dinosaur footprints from the Peace River Canyon, British Columbia, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology, 28:103-115.
Falkingham, P.L., Bates, K.T., Margetts, L., and Manning, P.L. 2011. The 'Goldilocks' effect: preservation bias in vertebrate track assemblages. Journal of the Royal Society Interface, 8:1142-1154.
Farlow, J.O., Pittman, J.G., and Hawthorne, J.M. 1989. Brontopodus birdi, Lower Cretaceous sauropods footprints from the U.S. Gulf Coastal Plain, p. 371-394. In Gillette, D.D. and Lockley, M.G. (eds.), Dinosaur tracks and traces. Cambridge University Press, Cambridge.
Fornós, J.J., Bromley, R.G., Clemmensen, L.B., and Rodriguez-Perea, A. 2002. Tracks and trackways of Myotragus balearicus Bate (Artiodactyla, Caprinae) in Pleistocene aeolianites from Mallorca (Balearic Islands, western Mediterranean). Palaeogeography,Palaeoclimatology, Palaeoecology, 180:277-313.
Gangloff, R.A., May, K.C., and Storer, J.E. 2004. An early Late Cretaceous dinosaur tracksite in Central Yukon Territory, Canada. Ichnos, 11:299-309.
Gatesy, S. 2003. Direct and indirect track features: what sediment did a dinosaur touch? Ichnos, 10:91-98.
Gatesy, S.M., Middleton, K.M., Jenkins Jr, F.A., and Shubin, N.H. 1999. Three-dimensional preservation of foot movements in Triassic theropod dinosaurs. Nature, 399:141-144.
Haubold, H. 1996. Ichnotaxonomie und Klassifikation von Tetrapodenfährten aus dem Perm. Hallesches Jahrbuch für Geowissenschaften, B 18:23-88.
Haubold, H. 1998. The Early Permian tetrapod ichnofauna of Tambach, the changing concepts in ichnotaxonomy. Hallesches Jahrbuch für Geowissenschaften, B 20:1-16.
Hornung, J.J., Böhme, A., van der Lubbe, T., Reich, M., and Richter, A. 2012. Vertebrate tracksites in the Obernkirchen Sandstone (late Berriasian, Early Cretaceous) of northwest Germany -- their stratigraphical, palaeogeographical, palaeoecological, and historical context. Paläontologische Zeitschrift, 86:231-267.
Hunt, A.P., Lockley, M.G., and Lucas, S.G. 1994. Trackway evidence for tetrapod predation from the Early Permian of southern New Mexico. New Mexico Geology, 16:84-85.
Hunt, A.P. and Lucas, S.G. 1998. Ichnological evidence for tetrapod predation in the Paleozoic: Is there any?, p. 59-62. In Lucas, S.G., Estep, J.W. and Hoffer, J.M. (eds.), Permian stratigraphy andpaleontology of the Robledo Mountains, NewMexico. New Mexico Museum of Natural History and Science Bulletin 12.
Kittl, E. 1891. Saurier Fährte von Bozen. Mitteilungen Österreichischen Touristen Klub, 3:7.
Klaus, W. 1963. Sporen aus dem südalpinen Perm. Jahrbuch der Geologischen Bundesanstalt, 106:229-361.
Kramer, J.M., Erickson, B.R., Lockley, M.G., Hunt, A.P., and Braddy, S.J. 1995. Pelycosaur predation in the Permian: Evidence from Laoporus trackways from the Coconino Sandstone with description of a new species of Permichnium, p. 245-249. In Lucas, S.G. and Heckert, A.B. (eds.), Early Permian Footprints and Facies . New Mexico Museum of Natural History and Science Bulletin 6.
Kustatscher, E., Bauer, K., Butzmann, R., Fischer, T.C., Meller, B., van Konijnenburg-van Cittert, J.H.A., and Kerp, H. 2014. Sphenophytes, pteridosperms and possible cycads from the Wuchiapingian (Lopingian, Permian) of Bletterbach (Dolomites, Northern Italy). Review ofPalaeobotany and Palynology, 208:65-82.
Kustatscher, E., van Konijnenburg-van Cittert, J.H.A., Bauer, K., Butzmann, R., Meller, B., and Fischer, T.C. 2012. A new flora from the Upper Permian of Bletterbach (Dolomites, N-Italy). Review of Palaeobotany and Palynology, 182:1-13.
Jackson, S.J., Whyte, M.A., and Romano, M. 2009. Laboratory-controlled simulations of dinosaur footprints in sand: a key to understanding vertebrate track formation and preservation. Palaios, 24:222-238.
Jackson, S.J., White, M.A., and Romano, M. 2010: Range of experimental dinosaur ( Hypsilophodonfoxii ) footprints due to variation in sand consistency: how wet was the track? Ichnos, 17:197- 214.
Latorre Macarrón, P., Pascual Arribas, C., Sanz Pérez, E., and Hernández Medrano, N. 2006. El yacimiento con huellas de saurópodos de Miraflores I, Fuentes de Magaña (Soria, España), p. 273-296. In Colectivo Arqueológico-Paleontológico Salense (ed.), Actas de las III Jornadas sobre Dinosaurios y su Entorno. Salas de los Infantes, Burgos, España.
Leonardi, G. 1987. Glossary and Manual of Tetrapod Footprint Palaeoichnology. Departamento Nacional da Produção Mineral, Brasilia, Brazil.
Leonardi, G. and Nicosia, U. 1973. Stegocephaloid footprint in the Middle Permian sandstone (Groedener Sandsteine) of the Western Dolomites. Annali dell’Università di Ferrara, NuovaSerie, 9:116-124.
Leonardi, P. 1948. Contributi alla conoscenza della flora delle Arenarie di Val Gardena (Permiano medio-inf.) dell’Alto Adige: la nuova flora di Redagno e una felce di Egna. Memoriedell’Istituto Geologico Mineralogico dell’Università di Padova, 16:3-15.
Leonardi, P. 1951. Orme di tetrapodi nelle Arenarie di Val Gardena (Permiano medio-inferiore) dell’Alto Adige sud-orientale. Memorie dell’Istituto Geologico Mineralogico dell’Università diPadova, 17:1-23.
Leonardi, P., Conti, M.A., Leonardi, G., Mariotti, N., and Nicosia, U. 1975. Pachypes dolomiticus n. gen. n. sp.; Pareiasaur footprint from the “Arenaria di Val Gardena” (Middle Permian) in the Western Dolomites (N. Italy). Accademia Nazionale dei Lincei, Rendiconti Classe Scienzematematiche fisiche e naturali, serie VIII, 57:221-232.
Lockley, M.G., Houck, K.J., and Prince, N.K. 1986. North America’s largest dinosaur trackway site: implications for Morrison Formation paleoecology. Geological Society of America Bulletin, 97:1163-1176.
Lockley, M.G. and Madsen, J.H. 1993. Early Permian vertebrate trackways from the CedarMesa Sandstone of eastern Utah: evidence of predator-prey interaction. Ichnos, 2:147-153.
Marty, D., Strasser, A., and Meyer, C.A. 2009. Formation and taphonomy of human footprints in microbial mats of present-day tidal-flat environments: implications for the study of fossil footprints. Ichnos, 16:127-142.
Massari F., Conti, M.A., Fontana, D., Helmod, K., Mariotti, N., Neri, C., Nicosia, U., Ori, G.G., Pasini, M., and Pittau, P. 1988. The Val Gardena Sandstone and Bellerophon Formation in the Bletterbach Gorge (Alto Adige, Italy): biostratigraphy and sedimentology. Memorie di Scienze Geologiche, 40:229-273.
Massari, F., Neri, C., Pittau, P., Fontana, D., and Stefani, C. 1994. Sedimentology, palynostratigraphy and sequence stratigraphy of a continental to shallow-marine rift-related succession: Upper Permian of the eastern Southern Alps (Italy). Memorie di ScienzeGeologiche, 46:119-243.
Matsukawa, M., Matsui, T., and Lockley, M.G. 2001. Trackway evidence of herd structure among ornithopod dinosaurs from the Cretaceous Dakota Group of northeastern New Mexico, USA. Ichnos, 8:197-206.
Melchor, R.N. and De Valais, S. 2006. A review of Triassic tetrapod track assemblages from Argentina. Palaeontology, 49:355-379.
Mezga, A. and Bajraktarevic, Z. 1999. Cenomanian dinosaur tracks on the islet of Fenoliga in southern Istria, Croatia. Cretaceous Research, 20:735-746.
Milàn, J. 2006. Variations in the morphology of the emu ( Dromaius novaehollandiae ) tracks reflecting differences in walking pattern and substrate consistency: ichnotaxonomic implications. Palaeontology, 49:405-420.
Milàn, J. and Bromley, R.G. 2006. True tracks, undertracks and eroded tracks, experimental work with tetrapod tracks in laboratory and field. Palaeogeography, Palaeoclimatology,Palaeoecology, 231:253-264.
Milàn, J. and Bromley, R.G. 2008. The impact of sediment consistency on track and undertrack morphology: experiments with emu tracks in layered cement. Ichnos, 15:19-27.
Milàn, J., Avanzini, M., Clemmensen, L.B., Garciá-Ramos, J.C., and Piñuela, L. 2006. Theropod foot movement recorded by Late Triassic, Early Jurassic and Late Jurassic fossil footprints, p. 352-364. In Harris et al. (eds.), The Triassic-Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin, 37.
Nadon, G.C. 2001. The impact of sedimentology on vertebrate track studies, p. 395-407. In Tanke, D.H. and Carpenter, K. (eds.), Mesozoic Vertebrate Life. Indiana University Press, Indiana.
Nicosia, U., Petti, F.M., Perugini, G., D’Orazi Porchetti, S., Sacchi, E., Conti, M.A., and Mariotti, N. 2007. Dinosaur tracks as paleogeographic constraints: new scenarios for the Cretaceous geography of the periadriatic region. Ichnos, 14:69-90.
Peabody, F.E. 1948. Reptile and Amphibian trackways from the Lower Triassic Moenkopi Formation of Arizona and Utah. Bulletin of the Department of Geological Sciences , Berkeleyand Los Angeles, 27:295-468.
Rainforth, E.C. 2005 . Ichnotaxonomy of the fossil footprints of the Connecticut Valley (early Jurassic, Newark Supergroup, Connecticut and Massachusetts). PhD Thesis, Columbia University, New York.
Razzolini, N.L., Vila, B., Castanera, D., Falkingham, P.L., Barco, J.L., Canudo, J.I., Manning, P.L., and Galobart, A. 2014. Intra-trackway morphological variations due to substrate consistency: the El Frontal Dinosaur Tracksite (Lower Cretaceous, Spain). PLoS ONE, 9(4):e93708.
doi:10.1371/journal.pone.0093708
Roghi, G., Kustatscher, E., Bernardi, M., Dal Corso, J., Forte, G., Franz, M., Hochuli, P., Krainer, K., Petti, F.M., Ragazzi, E., Riva, A., Wappler, T., and Gianolla, P. 2014. Field trip to Permo-Triassic palaeobotanical and palynological sites of the Southern Alps. Geo.Alp,Geologisch-Paläontologische Mitteilungen Innsbruck, 11:29-84.
Romano, M., Citton, P., and Nicosia, U. 2015. Corroborating trackmaker identification through footprint functional analysis: the case study of Ichniotherium and Dimetropus. Lethaia, 49:102-116.
Romero-Molina, M., Sarjeant, W., Perez-Lorente, F., Lopez, A., and Requeta, E. 2003. Orientation and characteristics of theropod trackways from the Las Losas palaeoichnological site (La Rioja, Spain). Ichnos, 10:241-254.
Seitz, S.M., Curless, B., Diebel, J., Scharstein, D., and Szeliski, R. 2006. A comparison and evaluation of multi-view stereo reconstruction algorithms. Proceedings of the 2006 IEEEComputer Society Conference on Computer Vision and Pattern Recognition(CVPR”06) 0-7695-2597-0/06.
Torcida Fernández-Baldor, F., Izquierdo Montero, L.A., Huerta, P., Montero Huerta, D. Pérez Martínez, G., and Urién Montero, V. 2006. El yacimiento de icnitas de dinosaurios de Costalomo (Salas de los Infantes, Burgos, España): nuevos datos, p. 313-347. In Colectivo Arqueológico-Paleontológico Salense (ed.), Actas de las III Jornadas sobre Dinosaurios y su Entorno. Salas de los Infantes, Burgos, España.
Ullman, S. 1979. The interpretation of structure from motion. Proceedings of the Royal Society ofLondon, B203:405-426.
Valentini, M., Conti, M.A., and Mariotti, N. 2007. Lacertoid footprints of the Upper Permian Arenaria di Val Gardena Formation (Northern Italy). Ichnos, 14:193-218.
Valentini, M., Conti, M.A., and Nicosia, U. 2008. Linking tetrapod tracks to the biodynamics, paleobiogeography, and paleobiology of their trackmakers: Pachypes dolomiticus Leonardi et al., 1975, a case study. Studi Trentini di Scienze NaturaliActa Geologica, 83:237-246.
Valentini, M., Nicosia, U., and Conti, M.A. 2009. A re-evalution of Pachypes, a pareiasaurian track from the Late Permian. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 251/1:71-94.
von Richthofen, F. 1860. Geognostische Beschreibung der Umgegend von Predazzo, S. Cassian und der Seisser Alpe in Südtirol. Perthes, Gotha.
Wilson, J.A., Marsicano, C.A., and Smith, R.M.H. 2009. Dynamic locomotor capabilities revealed by early dinosaur trackmakers from southern Africa. PLoS ONE 4(10):e7331.
doi:10.1371/journal.pone.0007331
Zhang, J., Daqing, L., Minglu, L., Lockley, M.G., and Bai, Z. 2006. Diverse dinosaur-, pterosaur-, and bird-track assemblages from the Hakou Formation, Lower Cretaceous of Gansu province, northwest China. Cretaceous Research, 27:44-55.
