

 Late Oligocene decapod crustaceans from the Trbovlje Formation of Slovenia, with a description of two new species of hymenosomatid crabs
Late Oligocene decapod crustaceans from the Trbovlje Formation of Slovenia, with a description of two new species of hymenosomatid crabs
Article number: 27.1.a4
https://doi.org/10.26879/1312
Copyright Paleontological Society, January 2024
Proceedings of the 8th Symposium on Fossil Decapod Crustaceans
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 10 June 2023. Acceptance: 1 December 2023.
ABSTRACT
From the Oligocene (Chattian) marls of the Trbovlje Formation, exposed at the Neža locality, Slovenia, a brachyuran crab (Malacostraca: Brachyura) association consisting of three taxa is described. The material presented here includes as many as 15 near-complete crab specimens from the Upper Oligocene of Slovenia does not only allow direct comparison with extant hymenosomatids, but the identification of important diagnostic characters permit classification of the fossils in question within extant genera, i.e., Halicarcinus White, 1846, and Lucascinus Poore, Guinot, Komai, and Naruse, 2016. The newly erected species, Halicarcinus popeius sp. nov. and Lucascinus trifailensis sp. nov., are considered herein the first reliable occurrences of the family Hymenosomatidae MacLeay, 1838, in the fossil record. Previously reported occurrence of hymenosomatid crabs from the Cretaceous of Brazil is disputed herein. The hymenosomatid association from the Neža locality is accompanied with the portunid crab, Necronectes cf. michelini A. Milne-Edwards, 1860, documented by an incomplete claw.
Rok Gašparič. Institute for Palaeobiology and Evolution, Novi trg 59, 1241 Kamnik, Slovenia and Oertijdmuseum, Bosscheweg 80, 5283 WB Boxtel, the Netherlands. rok.gasparic@gmail.com
Matúš Hyžný. Comenius University, Faculty of Natural Sciences, Department of Geology and Palaeontology, Bratislava, Slovakia. matus.hyzny@uniba.sk
Tomaž Hitij. Institute for Palaeobiology and Evolution, Novi trg 59, 1241 Kamnik, Slovenia and University of Ljubljana, Faculty of Medicine, Vrazov trg 2, 1000 Ljubljana, Slovenia. tomazhitij@gmail.com
Aleš Šoster. University of Ljubljana, Faculty of Natural Sciences and Engineering, Aškerčeva c. 12, SI-1000, Ljubljana, Slovenia. ales.soster@ntf.uni-lj.si
Key words: Chattian; Malacostraca; Brachyura; Hymenosomatidae; Portunidae Palaeoecology
Final citation: Gašparič, Rok, Hyžný, Matúš, Hitij, Tomaž, and Šoster, Aleš. 2024. Late Oligocene decapod crustaceans from the Trbovlje Formation of Slovenia, with a description of two new species of hymenosomatid crabs. Palaeontologia Electronica, 27(1):a4.
https://doi.org/10.26879/1312
palaeo-electronica.org/content/2024/5084-chattian-hymenosomatids
Copyright: January 2024 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
https://zoobank.org/71FEB770-6EE3-4621-B752-872A1CBDB605
INTRODUCTION
Representatives of the eubrachyuran crab family Hymenosomatidae MacLeay, 1838, are sometimes called “false spider crabs”, referring to their superficial resemblance to “true spider crabs” of the superfamily Majoidea Samouelle, 1819. In fact, the fossils of hymenosomatid crabs presented herein were first recognized as belonging to majoid spider crabs (Kovalchuk et al., 2023). Hymenosomatids are small eubrachyurans with only a lightly mineralised exoskeleton (Lucas, 1981), thus having a very low fossilization potential (Klompmaker et al., 2017, and references therein). Their relative absence in the fossil record was attributed to their small size, flat body, and poorly calcified carapace (Guinot, 2011, p. 46). This is likely connected to collection bias, rather than poor fossilization potential, as their remains are easily overlooked, due to their size. Because the recently published alleged occurrence of fossil hymenosomatid crabs from the Cretaceous of Brazil (Mendes et al., 2022) is disputed herein (see further below), the present occurrence from the Upper Oligocene of Slovenia is considered as the first and so far the only known report of fossil Hymenosomatidae.
Decapod occurrences in Oligocene and lower Miocene fish beds (i.e., laminated marls yielding exceptionally preserved articulated fish fauna) of the Central and primarily Eastern Paratethys have been recognized since the beginning of the twentieth century (Paucă, 1929; Smirnov, 1929). These occurrences have recently been reassessed by Hyžný et al. (2022) and Kovalchuk et al. (2023).
In the present contribution we describe well-preserved decapod specimens from the upper Oligocene fish beds of the Trbovlje Formation, Slovenia. Previously reported occurrence of decapods from the respective formation includes a single specimen of the xanthoid panopeid Glyphithyreus sulcatus (Beurlen, 1939) (Hyžný et al., 2020).
GEOLOGICAL SETTINGS
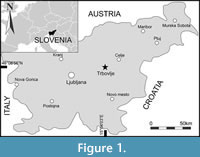 Decapod specimens were collected from the locality “Neža” (GPS coordinates: 46°08’56”N, 15°04’03”E) situated halfway between towns of Hrastnik and Trbovlje (Figure 1). Fossil-bearing Oligocene beds crop out in an abandoned coal mining pit, which has been restored and now represents an outdoor activity area. The wider area was intensively mined for lignite (brown coal) over the last two centuries and has been well researched (Bittner, 1884; Petrascheck, 1952; Kuščer, 1967; Jelen et al., 1992; Placer, 1999; Hafner, 2000). The Cenozoic sequence starts with the upper Oligocene Trbovlje Formation, which discontinuously overlies Triassic rocks. Basal horizons comprise conglomerates and sandstones, grading upwards into greyish coloured marls and marly limestones. An economically important coal seam occurs within the marly beds (Bechtel et al., 2004) and palynological analysis has identified species of Cupressaceae within the main coal seam (Bruch, 1998; Križnar, 2000) with a likely transition to reed marshes in the upper part. The marls and marly limestones above the coal layer are fossiliferous, yielding a diverse fish fauna (Bittner, 1884; Križnar, 2015; Križnar and Lorencon, 2021), crustaceans (Buckeridge et al., 2020; Kovalchuk et al., 2023; herein), molluscs (Bittner, 1884), as well as abundant plant remains (Lorencon, 2019). A late Oligocene (Chattian) age has been confirmed by radiometric dating of interbedded tuff layers (Odin et al., 1994). A grey marine clay of the Sivica Formation occurs on top of the fossiliferous beds; above this the sequence grades into lenses of fine-grained clastic rocks, particularly sandstones and conglomerates of the lower Miocene Govce Formation (Hafner, 2000).
Decapod specimens were collected from the locality “Neža” (GPS coordinates: 46°08’56”N, 15°04’03”E) situated halfway between towns of Hrastnik and Trbovlje (Figure 1). Fossil-bearing Oligocene beds crop out in an abandoned coal mining pit, which has been restored and now represents an outdoor activity area. The wider area was intensively mined for lignite (brown coal) over the last two centuries and has been well researched (Bittner, 1884; Petrascheck, 1952; Kuščer, 1967; Jelen et al., 1992; Placer, 1999; Hafner, 2000). The Cenozoic sequence starts with the upper Oligocene Trbovlje Formation, which discontinuously overlies Triassic rocks. Basal horizons comprise conglomerates and sandstones, grading upwards into greyish coloured marls and marly limestones. An economically important coal seam occurs within the marly beds (Bechtel et al., 2004) and palynological analysis has identified species of Cupressaceae within the main coal seam (Bruch, 1998; Križnar, 2000) with a likely transition to reed marshes in the upper part. The marls and marly limestones above the coal layer are fossiliferous, yielding a diverse fish fauna (Bittner, 1884; Križnar, 2015; Križnar and Lorencon, 2021), crustaceans (Buckeridge et al., 2020; Kovalchuk et al., 2023; herein), molluscs (Bittner, 1884), as well as abundant plant remains (Lorencon, 2019). A late Oligocene (Chattian) age has been confirmed by radiometric dating of interbedded tuff layers (Odin et al., 1994). A grey marine clay of the Sivica Formation occurs on top of the fossiliferous beds; above this the sequence grades into lenses of fine-grained clastic rocks, particularly sandstones and conglomerates of the lower Miocene Govce Formation (Hafner, 2000).
MATERIAL AND METHODS
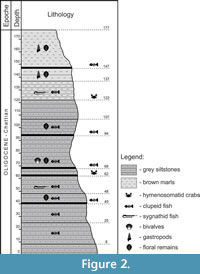 Due to the abundance of fossil fish remains, the “Neža” locality was frequently visited by private collectors in the recent years, resulting in discovery of several decapod specimens. Therefore, a detailed bed by bed sampling was done within the available 2m thick sequence. An area of approximately 1 m2 was excavated and all fossils on the beds were documented. Fossil remains of fish and flora occur throughout the section with varying frequency. Additionally, four mass accumulations of fish remains (probably a mass mortality event) were identified at different levels within the section, as well as two beds from which the presented decapod specimens originated (Figure 2).
Due to the abundance of fossil fish remains, the “Neža” locality was frequently visited by private collectors in the recent years, resulting in discovery of several decapod specimens. Therefore, a detailed bed by bed sampling was done within the available 2m thick sequence. An area of approximately 1 m2 was excavated and all fossils on the beds were documented. Fossil remains of fish and flora occur throughout the section with varying frequency. Additionally, four mass accumulations of fish remains (probably a mass mortality event) were identified at different levels within the section, as well as two beds from which the presented decapod specimens originated (Figure 2).
The present study is based on 15 specimens of near-complete crabs and a single large claw. The specimens were prepared and studied under a Leica EZ 4D stereomicroscope. Photographs were taken with a digital camera Nikon Zfc equipped with Micro NIKKOR 40mm f/2.8G lens. Photographs were subsequently developed in Photoshop CS6 to correct for light, contrast, and colour where needed. The large claw specimen was further prepared as a latex cast and whitened with sublimate of ammonium chloride prior to photography. Selected near-complete specimens were documented under SEM ThermoFischer Scientific Quattro S. Micrographs were acquired in low vacuum mode with an accelerating voltage of 15 kV and a spot size of 3.5 in backscattered electron imaging mode using the annular CBS detector. Samples were coated with a thin film of amorphous carbon to ensure electrical conductivity. Measurements of the specimens were made with digital callipers under the stereomicroscope. The reference of size is given as maximum carapace width, which is mostly at the level of the posterior part of carapace. The rostrum is included in measurements of carapace length. Walking leg length is measured from the longest leg, usually the second pereopod (P2).
Abbreviations
Abbreviations of dorsal carapace characters used in the illustrations are as follows: CW - Carapace width, CL - Carapace length, P1L - Cheliped length, P2L - Pereopod P2 length. Repository acronyms: MNHN.F - Collection de Paléontologie, Muséum national dʼHistoire naturelle, Paris, France. RGA/SMNH - Slovenian Museum of Natural History, Ljubljana, Slovenia (R. Gašparič Collection).
SYSTEMATIC PALAEONTOLOGY
Order DECAPODA Latreille, 1802
Infraorder BRACHYURA Latreille, 1802
Superfamily HYMENOSOMATOIDEA MacLeay, 1838
Family HYMENOSOMATIDAE MacLeay, 1838
Genus HALICARCINUS White, 1846
Type species. Cancer planatus Fabricius, 1775, by original designation.
Remarks. Hymenosomatidae MacLeay, 1838, comprises 24 genera and over 130 species (Poore et al., 2016) of which Halicarcinus White, 1846, was the most diverse and morphologically heterogeneous. Poore et al. (2016) have revised Halicarcinus and reassigned the species to seven genera to accommodate the diversity previously encompassed in the genus. Currently Halicarcinus includes seven species characterised by the complete hymenosomian groove, well-developed epistome, rostrum developed as median tubercle with paired pseudorostral elements, strongly curved gonopod 1, pleomeres 1-5 free, pleomere 6 fused to telson (pleotelson), pereopodal dactylar teeth shorter than in other genera, and propodi of male chelipeds frequently swollen and barrel-like (Naruse and Komai, 2009; Poore et al., 2016).
Halicarcinus popeius sp. nov.
Figure 3, Figure 4A-D, Figure 5A-C
zoobank.org/BC3F122F-89A8-4BDB-A174-664B834FA1D1
2023 Majoidea gen. et sp. indet. - Kovalchuk et al., p. 5, figs. 2E, F.
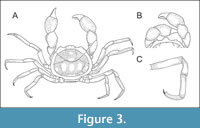 Etymology. The species name originates from a Latinised name of the cartoon character Popeye, the sailor man. A fictional character who, despite the diminutive stature, possesses strong, bulging forearms, reminiscent of the chelipeds of the new species.
Etymology. The species name originates from a Latinised name of the cartoon character Popeye, the sailor man. A fictional character who, despite the diminutive stature, possesses strong, bulging forearms, reminiscent of the chelipeds of the new species.
 Diagnosis. Carapace oval, wider than long, surrounded by a wide rim. Trilobate to trapezoid rostrum; length not extending as far as eyes. Eyes on short, stout eyestalks; extending beyond rostrum. Anterolateral corners forming a rounded postocular projection; either as rounded lobe or a pair of forward pointing teeth. Lateral side of carapace rounded; almost vertical posteriorly; with a concavity at posterolateral margin. Dorsal carapace with continuous gastro-cardiac groove, forming a hexagonal cardiac region. Chelipeds massive in males, sub-equal, homomorph, with swollen barrel-like propodus; more slender in females. Both fingers long, occluding with a gape; no dentation of the occlusal margin. Pereopods very long and slender. Dactylus broad, distinctively curved, tip composed of two developed, sub-equal, teeth; ventral edge armed by a row of blunt tubercles.
Diagnosis. Carapace oval, wider than long, surrounded by a wide rim. Trilobate to trapezoid rostrum; length not extending as far as eyes. Eyes on short, stout eyestalks; extending beyond rostrum. Anterolateral corners forming a rounded postocular projection; either as rounded lobe or a pair of forward pointing teeth. Lateral side of carapace rounded; almost vertical posteriorly; with a concavity at posterolateral margin. Dorsal carapace with continuous gastro-cardiac groove, forming a hexagonal cardiac region. Chelipeds massive in males, sub-equal, homomorph, with swollen barrel-like propodus; more slender in females. Both fingers long, occluding with a gape; no dentation of the occlusal margin. Pereopods very long and slender. Dactylus broad, distinctively curved, tip composed of two developed, sub-equal, teeth; ventral edge armed by a row of blunt tubercles.
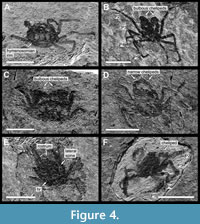
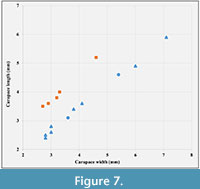
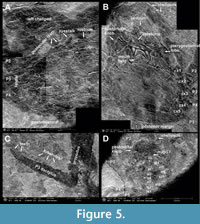 Type material. Holotype is RGA/SMNH 6377-6378 (part and counterpart) (Figure 4A), paratypes are RGA/SMNH 6373-6374 (part and counterpart) (Figure 4B, Figure 6C), RGA/SMNH 6371-6372 (part and counterpart), RGA/SMNH 6369-6370 (part and counterpart), RGA/SMNH 6385-6386 (part and counterpart) (Figure 4C, Figure 5A) and RGA/SMNH 2041 (Figure 4D, Figure 5B). Other specimens are RGA/SMNH 2039-2040 (part and counterpart), RGA/SMNH 6375-6376 (part and counterpart), RGA/SMNH 6381-6382 (part and counterpart) and RGA/SMNH 6389-6390 (part and counterpart). Measurements are given in Figure 7, Table 1 and Table 2.
Type material. Holotype is RGA/SMNH 6377-6378 (part and counterpart) (Figure 4A), paratypes are RGA/SMNH 6373-6374 (part and counterpart) (Figure 4B, Figure 6C), RGA/SMNH 6371-6372 (part and counterpart), RGA/SMNH 6369-6370 (part and counterpart), RGA/SMNH 6385-6386 (part and counterpart) (Figure 4C, Figure 5A) and RGA/SMNH 2041 (Figure 4D, Figure 5B). Other specimens are RGA/SMNH 2039-2040 (part and counterpart), RGA/SMNH 6375-6376 (part and counterpart), RGA/SMNH 6381-6382 (part and counterpart) and RGA/SMNH 6389-6390 (part and counterpart). Measurements are given in Figure 7, Table 1 and Table 2.
Type locality. Trbovlje, Slovenia.
 Description. Carapace oval, wider than long (CL/CW = 0.82-0.93) (Table 1), surrounded by a wide hymenosomian rim. Trilobate to trapezoid rostrum; wide and short; width approx. 0.1-0.2 times carapace width; length not extending as far as eyes (Figure 5A). Lobes close together; sub equal in length; median lobe slightly bigger than the other two, sometimes not differentiated and forming a single trapezoidal rostrum (Figure 4B). Eyes on short, stout eyestalks; extending beyond rostrum (Figure 5A). Anterolateral corners rounded, forming a rounded postocular projection; either as rounded lobe or a pair of forward pointing teeth. Lateral side of carapace rounded; almost vertical posteriorly; with a concavity at posterolateral corner (Figure 4A-D).
Description. Carapace oval, wider than long (CL/CW = 0.82-0.93) (Table 1), surrounded by a wide hymenosomian rim. Trilobate to trapezoid rostrum; wide and short; width approx. 0.1-0.2 times carapace width; length not extending as far as eyes (Figure 5A). Lobes close together; sub equal in length; median lobe slightly bigger than the other two, sometimes not differentiated and forming a single trapezoidal rostrum (Figure 4B). Eyes on short, stout eyestalks; extending beyond rostrum (Figure 5A). Anterolateral corners rounded, forming a rounded postocular projection; either as rounded lobe or a pair of forward pointing teeth. Lateral side of carapace rounded; almost vertical posteriorly; with a concavity at posterolateral corner (Figure 4A-D).
 Dorsal carapace with continuous gastro-cardiac groove, extending concavely from one side of hymenosomian rim to the other; forming a hexagonal cardiac region (Figure 4A, D). Frontal region trapezoidal; not differentiated from gastric region posteriorly. Hepatic and prebranchial regions small; triangular. Postbranchial region square; separated from cardiac region by the thoracic groove. Cardiac region ornamented by three longitudinally oval areas of brighter colour (Figure 4A); smaller oval coloration of dorsal carapace also in gastric region, anteriorly of gastro-cardiac groove; crescent-shaped coloration of carapace at posterior border.
Dorsal carapace with continuous gastro-cardiac groove, extending concavely from one side of hymenosomian rim to the other; forming a hexagonal cardiac region (Figure 4A, D). Frontal region trapezoidal; not differentiated from gastric region posteriorly. Hepatic and prebranchial regions small; triangular. Postbranchial region square; separated from cardiac region by the thoracic groove. Cardiac region ornamented by three longitudinally oval areas of brighter colour (Figure 4A); smaller oval coloration of dorsal carapace also in gastric region, anteriorly of gastro-cardiac groove; crescent-shaped coloration of carapace at posterior border.
Chelipeds massive, with swollen barrel-like propodus (Figure 4A, C); length approx. 1.1-1.7 times of carapace length; slighter in females. Ischium short; merus slightly longer than wide; carpus longer than merus, swollen with rounded margins, almost as broad as long; manus greatly inflated, margins rounded and convex. Both fingers long, occluding with a gape; no dentation of the occlusal margin observable. Pereopods very long and slender (Figure 4B, Table 2), almost same length, approx. 2.0-3.5 times of carapace length. Ischium very short; merus longest; carpus about one-thirds as long as merus; propodus almost as long as merus. Dactylus shorter than the propodus, broad, distinctively curved, tip composed of two developed, sub-equal, teeth; ventral edge armed by a row of blunt tubercles (Figure 5C).
Remarks. Halicarcinus popeius sp.nov. is the first fossil representative of the genus. Thus, by the general habitus it would be most similar to Halicarcinus quoyi (H. Milne Edwards, 1853), with which it shares a wider than long carapace with broad rim, a trilobed rostrum, shape of pereopodal dactyli and strongly swollen, bulbous propodi of male chelipeds. The new fossil species differs from H. quoyi in possessing postorbital projections at anterolateral corners of carapace and in development of dorsal carapace regions, especially wide hexagonal cardiac region. Additionally, we recognize heterogeneity in development of rostra within the fossil material, which can be explained as intraspecific variations or possible sexual dimorphism. Lucas (1981) previously recognized that the shape and relative length of the rostrum within Halicarcinus are sometimes sexually dimorphic, although such variations were not reported in Halicarcinus quoyi.
Range. Late Oligocene (Chattian).
Occurrence. To date, known only from the type locality.
Genus LUCASCINUS
Poore, Guinot, Komai, and Naruse, 2016
Type species. Halicarcinus bedfordi Montgomery, 1931, by original designation.
Remarks. Poore et al. (2016) erected a genus Lucascinus and transferred to it three species: Lucascinus bedfordi (Montgomery, 1931) (originally as Halicarcinus); Lucascinus coralicola (Rathbun, 1909) (originally as Rhynchoplax) and Lucascinus keijibabai (Takeda and Miyake, 1971) (originally as Rhynchoplax). The species within Lucascinus differ from Rhynchoplax mostly by possessing a male pleon with free somites 1-5, plus telson, and additionally from Halicarcinus by their elongated chelipeds and triangular plate-like rostrum (Poore et al., 2016).
Lucascinus trifailensis sp. nov.
Figure 4E-F, Figure 5D, Figure 6
zoobank.org/95CB277A-BA14-4912-999F-8C69D499D24E
Etymology. The species name refers to old name for coal mining town of Trbovlje (Trifail). Trbovlje is Slovenia’s tenth-largest town and closest to the type locality of new species.
Diagnosis. Carapace trapezoid, longer than wide, fully surrounded by a rim. Rostrum formed as a long triangular plate. Postocular anterior margin sinuous; defined by a postocular tubercle and a more pronounced tooth at anterolateral corner of the carapace. Lateral side of carapace almost straight; diverging posteriorly; with a single projection near base of cheliped. Posterior border of carapace sinuously rounded. Dorsal carapace with visible gastro-cardiac groove and posteriorly developed thoracic grooves. Frontal region trapezoidal; not differentiated from gastric posteriorly. Cardiac region simple and posteriorly not bordered. Male pleon triangular; pleonal somites 1-5 free; somite 1 widest. Chelipeds long; propodus elongated and slightly swollen. Fingers short. Pereopods very long and slender; almost same length. Dactylus short, curved, teeth not observed.
Type material. Holotype is RGA/SMNH 6384-6383 (part and counterpart) (Figure 4E); paratypes are RGA/SMNH 6391 (Figure 4F), RGA/SMNH 6387-6388 (part and counterpart) (Figure 5D), RGA/SMNH 6379-6380 (part and counterpart) and RGA/SMNH 6392. Measurements are given in Figure 7, Table 1 and Table 2.
Type locality. Trbovlje, Slovenia.
Description. Carapace trapezoidal, longer than wide (CL/CW = 1.13-1.30) (Table 1), fully surrounded by a hymenosomian rim (Figure 4E - F). Rostrum formed as a long triangular plate; twice as long as wide; width approx. 0.1-0.15 times carapace width (Figure 5D). Postocular anterior margin sinuous; defined by a postocular tubercle and a more pronounced tooth at anterolateral corner of the carapace. Lateral side of carapace slightly convex; almost straight; diverging posteriorly; with a single projection near base of cheliped. Posterior border of carapace sinuously rounded (Figure 4F).
Dorsal carapace with visible gastro-cardiac groove and posteriorly developed thoracic grooves. Frontal region trapezoid; not differentiated from gastric posteriorly. Cardiac region simple and posteriorly not bordered (Figure 4E). Male pleon triangular; pleonal somites free; somite 1 widest, with rounded convex lateral sides; rest of the somites with straight and anteriorly converging lateral margins; pleotelson much longer than wide, triangular, with rounded apex (Figure 5D).
Chelipeds long; length approx. 0.85-1.0 times of carapace length. Ischium short; merus and carpus longer than wide; propodus elongated and lightly swollen. Fingers shorter than manus. Pereopods very long and slender (Figure 4F, Table 2); length approx. 1.9-2.3 times of carapace length, P3 and P4 longest. Ischium short; merus almost as long as propodus, carpus less than one third of merus; propodus longest, but narrower than the merus. Dactylus short, curved, teeth not observed.
Remarks. Lucascinus trifailensis sp. nov. is the first fossil representative of the genus and among extant species most closely resembles Lucascinus bedfordi (Montgomery, 1931). The shared similarities between the fossil and extant species are in ornamentation of dorsal carapace and the shape of a triangular rostrum. However, Lucascinus trifailensis sp. nov. has a narrower carapace, with distinct sinuous shape of the anterior margin, defined by a small postocular tubercle, and a more pronounced tooth at the anterolateral corner of the carapace. Additional spinose projection is located at lateral margin, near the base of cheliped, which is typical for Lucascinus (Poore et al., 2016), but much less pronounced in Lucascinus trifailensis sp. nov. Such combination of characters has not been observed in any of the extant species. However, we prefer to use the most parsimonious way and include the fossil species in the closest extant genus.
Range. Late Oligocene (Chattian).
Occurrence. To date, known only from the type locality.
Superfamily PORTUNOIDEA Rafinesque, 1815
Family PORTUNIDAE Rafinesque, 1815
Genus NECRONECTES A. Milne-Edwards, 1881
Type species. Necronectes vidalianus A. Milne-Edwards, 1881, by original designation.
Necronectes cf. michelini A. Milne-Edwards, 1860
Figure 8A-D
Material. A single “crusher” chela (RGA/SMNH 5009-5010) preserved as an imprint. Measurements: length of propodus = 53.6 mm; maximum height of manus = 31.9 mm; length of dactylus = 31.0 mm.
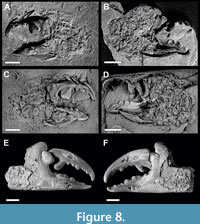 Description. Chela robust. Manus trapezoidal to suboval in outline; upper and lower margin arched and converging proximally; surface appears smooth, without indication of spines on distal margin at dactylus articulation. Fingers long and stout; as long as manus. Fixed finger robust; occlusal margin with four large molariform teeth, followed by several smaller conical teeth distally; first and second proximal teeth joined; fingertip curved upwards. Dactylus stout, curved forward; with rounded upper margin; strong proximal knob-like flattened molariform tooth followed by six rounded conical teeth of different sizes; strongly curved fingertip.
Description. Chela robust. Manus trapezoidal to suboval in outline; upper and lower margin arched and converging proximally; surface appears smooth, without indication of spines on distal margin at dactylus articulation. Fingers long and stout; as long as manus. Fixed finger robust; occlusal margin with four large molariform teeth, followed by several smaller conical teeth distally; first and second proximal teeth joined; fingertip curved upwards. Dactylus stout, curved forward; with rounded upper margin; strong proximal knob-like flattened molariform tooth followed by six rounded conical teeth of different sizes; strongly curved fingertip.
Remarks. The original cuticle has been dissolved and the massive fingers are preserved as hollow imprints. Therefore, some details cannot be observed in the studied specimen. However, after preparing the latex cast, the preserved features of the chela are sufficient to assign the material to Necronectes A. Milne-Edwards, 1881. Observed important taxonomic characters include lack of distal spines on manus at position of dactylus articulation and rounded upper margin of manus, which are the key characters for an assignment to Necronectes, and differentiate it from closely related Scylla De Haan, 1833 (Ossó and Gagnaison, 2019). The herein presented specimen strongly resembles Necronectes michelini Milne-Edwards, 1860, from the Miocene of France (Figure 8E-F), somewhat differing in well-developed proximal molariform teeth on fixed finger and dactylus. Given similarities in preserved specimen, the stratigraphic span and geographic distribution of N. michelini, it is also possible that the occurrence reported here, from the upper Oligocene of Slovenia, belongs to this species. Nevertheless, we refrain to assign it to N. michelini because of insufficient preservation of diagnostic features on the manus and absence preserved carapace remains. Due to the fragmentary preservation we leave the specimen in open nomenclature.
Range. Late Oligocene (Chattian).
Occurrence. Necronectes is a well-known genus known from a number of Oligocene and Miocene occurrences (Schweitzer et al., 2006; Luque et al., 2017; Hyžný and Dulai, 2021). Necronectes michelini A. Milne-Edwards, 1860, has so far been reported from the middle Miocene of France (Ossó and Gagnaison, 2019) and the middle Miocene of Hungary (Lőrenthey and Beurlen, 1929).
DISCUSSION
Fossil Hymenosomatidae
The present occurrence of hymenosomatid crabs from the upper Oligocene of Slovenia is considered the first reliable report of the respective family in the fossil record. Mendes et al. (2022) reported several small, incomplete, distorted dorsal carapaces from the Lower Cretaceous of NE Brazil and assigned them to three newly erected species within two newly erected genera belonging to a newly erected subfamily Eureotropisinae within the family Hymenosomatidae. Unfortunately, the preservation of their material lacking diagnostic synapomorphies of the family Hymenosomatidae does not warrant the attribution of the Brazilian material to the respective family. The carapaces of Mendes et al. (2022) are relatively featureless, and their comparison with any extant brachyuran higher taxa is difficult at best. Moreover, the longitudinal dorsal ridge of Eureotropisinae crossing the entire or nearly entire carapace as shown by Mendes et al. (2022) is not present in any known brachyuran taxon (Ng et al., 2008; Davie et al., 2015). Thus, the affinities of the Eureotropisinae remain obscure.
Taphonomy
Despite the relatively low fossilization potential of hymenosomatid crabs, due to their small size and weakly mineralized cuticle, the studied specimens from the Trbovlje Formation are always preserved fully articulated but compressed. The assessment of taxonomical characters is further complicated by what appears to be a thin layer of carbonized film covering the carapace obscuring the details, and in some cases even the exact outline of the carapace. This can possibly be contributed to the carbonization of the biofilm layer and organic matter that covered the dorsal carapace and camouflaged the hymenosomatids in their environment (Melrose, 1975).
The complete and articulated mode of preservation of hymenosomatid crabs indicates, that they likely represent corpses or a mix of corpses and moults (compare with Hyžný et al., 2022). Specimens would have been quickly buried in-situ, without any or a minimum post-mortem transportation or disturbance, and under calm, but rapid depositional conditions (Klompmaker et al., 2017).
The claw of a portunid crab Necronectes sp. is disarticulated, with pereopod fragments observed close by, which suggests that it is parautochthonous or even allochthonous, and was transported before burial by mild currents.
Palaeoenvironment
Extant hymenosomatid crabs are ecologically very diverse, inhabiting marine, estuarine, and even freshwater environments (Melrose, 1975; Guinot and Richer de Forges, 1997; Guinot and Mazincourt, 2020). Oligocene fish beds of the Paratethys are interpreted as deposited in various depositional settings, from deeper marine, with anoxic bottom conditions, and fast sedimentation rates, to possibly shallower costal settings or open lagoonal settings (Kovalchuk et al., 2023).
We conclude, that the late Oligocene fauna from the Neža locality inhabited a shallow marine, near shore facies. High influx of organic detritus based on rich plant remains, but calm environment without strong currents that would significantly disturb post-mortem taphocoenosis. The shallow depth is supported by the present ichthyofauna, which apart from predominant specimens of clupeid fish, consists of sygnathid and centriscid species having a bathymetrical preference for shallow near-shore bottoms with seagrass meadows (Ahnesjö and Craig, 2011; Nelson et al., 2016). Such interpretation of palaeoenvironment fits well with the presence of hymenosomatid crabs, as most extant species inhabit shallow water environment.
CONCLUSIONS
Two newly described species of the family Hymenosomatidae represent the first reliable occurrence of the family in the fossil record. Halicarcinus popeius sp. nov. and Lucascinus trifailensis sp. nov. are the first fossil representatives of the genera Halicarcinus and Lucascinus. The poor fossil record of the group is partly due to the fragile nature of hymenosomatid exoskeleton and their poor preservation potential, due to the lack of fossiliferous localities with calm depositional near shore facies, and because of the minute size of specimens also due to the presence of collecting bias. Shallow water, near shore environmental setting, similar as is inhabited by most extant hymenosomatid species, is reconstructed based on associated fauna.
ACKNOWLEDGEMENTS
We wish to thank R. Lorencon and M. Križnar for a company and help during fieldwork, A.Ž. Gašparič for her thorough proofreading and the journal reviewers, J. Luque, (Cambridge University) and two anonymous reviewers, for constructive comments on an earlier version of the manuscript. The research of MH has been supported by the Slovak Research and Development Agency under contracts no. APVV-20-0079 and APVV-22-0523.
References
Ahnesjö, I. and Craig, J.F. 2011. The biology of Syngnathidae: pipefishes, seadragons and seahorses. Journal of Fish Biology, 71:1597–1602.
https://doi.org/10.1111/j.1095-8649.2011.03008.x
Bechtel, A., Markic, M., Sachsenhofer, R.F., Jelen, B., Gratzer, R., Lücke, A., and Püttmann, W. 2004. Paleoenvironment of the upper Oligocene Trbovlje coal seam (Slovenia). International Journal of Coal Geology, 57:23–48.
https://doi.org/10.1016/j.coal.2003.08.005
Beurlen, K. 1939. Neue Dekapoden-Krebse aus dem ungarischen Tertiär. Paläontologische Zeitschrift, 21:135–161.
Bittner, A. 1884. Die Tertiär-Ablagerungen von Trifail und Sagor. Jahrbuch der kaiserlich-königlichen Geologischen Reichsanstalt, 34:236–281.
Bruch, A. 1998. Palynologische Untersuchungen im Oligozän Sloweniens - Paläo-Umwelt und Paläoklima im Ostalpenraum. Tübinger Mikropaläontologische Mitteilungen, 18:1–193.
Buckeridge, J.S., Kočí, T., Gašparič, R., and Kočová, V.M. 2020. Actinobalanus? sloveniensis (Thoracica, Balanoidea), a new species of cirripede from the Oligocene and Miocene of Slovenia that grew attached to wood substrates. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 296:51–65.
https://doi.org/10.1127/njgpa/2020/0902
Davie, P.J.F., Guinot, D., and Ng, P.K.L. 2015. Chapter 71-2. Anatomy and functional morphology of Brachyura. In Castro, P., Davie, P.J.F., Guinot, D., Schram, F.R., and von Vaupel Klein, J.C. (eds.), Treatise on Zoology – Anatomy, Taxonomy, Biology. The Crustacea, Part 9C-I:11–163.
https://doi.org/10.1163/9789004190832_004
de Haan, W. 1833-1850. Crustacea. In Von Siebold, P.F. (ed.), Fauna Japonica. Muller, Amsterdam, pp. xvii, xxxi, 244 pp.
Fabricius, J.C. 1775. Systema Entomologiae, sistens Insectorum Classes, Ordines, Genera, Species, adjectis Synonymis, Locis, Descriptionibus, Observationibus. Kortii, Flensburgi et Lipsiae, xxxii + 832 pp.
Guinot, D. 2011. The position of the Hymenosomatidae MacLeay, 1838, within the Brachyura (Crustacea, Decapoda). Zootaxa, 2890:40–52.
https://doi.org/10.11646/zootaxa.2890.1.4
Guinot, D. and Richer de Forges, B. 1997. Affinités entres les Hymenosomatidae MacLeay, 1838 et les Inachoididae Dana, 1851 (Crustacea, Decapoda, Brachyura). Zoosystema, 19:453–502.
Guinot, D. and de Mazincourt, V. 2020. A new freshwater crab of the family Hymenosomatidae MacLeay, 1838 from New Caledonia (Crustacea, Decapoda, Brachyura) and an updated review of the hymenosomatid fauna of New Caledonia. European Journal of Taxonomy, 671:1–29.
https://doi.org/10.5852/ejt.2020.671
Hafner, G. 2000. Sedimentološke značilnosti trboveljskih plasti. Unpublished MSc thesis, University of Ljubljana, 105 pp.
Hyžný, M., Gašparič, R., and Dulai, A. 2020. Revision of species Plagiolophus sulcatus Beurlen, 1939 (Decapoda, Brachyura) from the Oligocene of Hungary and Slovenia. Geologija, 63:83–91.
https://doi.org/10.5474/geologija.2020.009
Hyžný, M. and Dulai, A. 2021. Badenian Decapods of Hungary. GeoLitera, Institute of Geosciences, University of Szeged, Hungary, 291 pp.
Hyžný, M., Kovalchuk, O., Świdnicka, E., Berezovsky, A., Dumitriu, S., Grădianu, I., Stefaniak, K., and Barkaszi, Z. 2022. Revisiting brachyuran crabs (Malacostraca: Decapoda) from Oligocene and Miocene fish beds of Europe. Geologica Carpathica, 73:579–597.
https://doi.org/10.31577/GeolCarp.73.6.3
Jelen, B., Aničič, B., Brezigar, A., Buser, S., Cimerman, F., Drobne, K., Monostori, M., Kedves, M., Pavšič, J., and Skaberne, D. 1992. Model of positional relationships for Upper Paleogene and Miocene strata in Slovenia. Interdisciplinary Geological Conference on the Miocene Epoch, Abstracts and Field Trips, Ancona:71–72.
Klompmaker, A.A., Portell, R.W., and Frick, M.G. 2017. Comparative experimental taphonomy of eight marine arthropods indicates distinct differences in preservation potential. Palaeontology, 60:1–22.
https://doi.org/10.1111/pala.12314
Kovalchuk, O., Hyžný, M., Świdnicka, E., Barkaszi, Z., Berezovsky, A., Dumitriu, S., Grădianu, I., Gašparič, R., Přikryl, T., and Stefaniak, K. 2023. Taphonomy and palaeoecology of decapod crustaceans from Oligocene and Early Miocene fish beds of the Central and Eastern Paratethys. Historical Biology, 35(12):2253–2270.
https://doi.org/10.1080/08912963.2022.2127097
Križnar, M. 2000. Ksilotomske analize rjavih premogov iz Zasavja. Unpublished BSc thesis, University of Ljubljana, 62 pp.
Križnar, M. 2015. Oligocenska ihtiofavna (Teleostei) Slovenije - pregled in problematika najdišč. Geološki zbornik, 23:99–104.
Križnar, M. and Lorencon, R. 2021. Zgornjeoligocenske morske britvice (Centriscidae, Gasterosteiformes) iz Trbovelj (Zasavje, Slovenija): preliminarna raziskava. Geološki zbornik, 26:58–61.
Kuščer, D. 1967. Zagorski terciar (Tertiary formations of Zagorje). Geologija, 10:5–85.
Latreille, P.A. 1802. Histoire Naturelle, Générale et Particulière des Crustacés et des Insectes. Ouvrage Faisant suite à l'Histoire Naturelle Générale et Particulière, Composée par LeClerc de Buffon, et Rédigée par C.S. Sonnini, Membre de Plusieurs Sociétés Savantes. Dufart: Paris. Vol. 3. 468 pp.
Lorencon, R. 2019. Oligocenska flora v okolici Trbovelj. Konkrecija, 8:12–18.
Lőrenthey, E. and Beurlen, K. 1929. Die fossilen Decapoden der Lander der Ungarischen Krone. Geologica Hungarica, Series Palaeontologica, 3:1–420.
Lucas, J.S. 1981. Spider crabs of the family Hymenosomatidae (Crustacea; Brachyura) with particular reference to Australian species: systematics and biology. Records of the Australian Museum, 33:148–247.
https://doi.org/10.3853/j.0067-1975.33.1980.199
Luque, J., Schweitzer, C.E., Santana, W., Portell, R.W., Vega, F.J., and Klompmaker, A.A. 2017. Checklist of fossil decapod crustaceans from tropical America. part I: Anomura and Brachyura. Nauplius, 25(0):1–85.
https://doi.org/10.1590/2358-2936e2017025
MacLeay, W.S. 1838. Invertebratae. Vol. 2. In: Smith, A. Illustrations of the Zoology of South Africa. Elder Smith, London.
Melrose, M.J. 1975. The marine fauna of New Zealand: family Hymenosomatidae (Crustacea, Decapoda, Brachyura). New Zealand Oceanographic Institute Memoirs, 34:1–123.
Mendes, I.D., Santana, W., and Carvalho, I.S. 2022. Fossil hymenosomatid crabs (Crustacea: Decapoda) from the Lower Cretaceous of NE Brazil. Cretaceous Research, 134:105174.
https://doi.org/10.1016/j.cretres.2022.105174
Milne-Edwards, A. 1881. Note sur quelques Crustacés fossiles des environs de Biarritz. Annales des Sciences géologiques, 11(2):1–8.
Milne Edwards, H. 1853. Mémoire sur la famille des Ocypodiens. Annales des Sciences Naturelles (ser. 3), 20:163–228.
Montgomery, S.K. 1931. Report on the Crustacea Brachyura of the Percy Sladen Trust expedition to the Abrolhos Islands under the leadership of Prof. W.J. Dakin in 1913, along with other crabs from Western Australia. Journal of the Linnean Society of London (Zoology), 37:405–464.
Naruse, T. and Komai, T. 2009. A new species of the false spider crab genus Halicarcinus (Decapoda: Brachyura: Hymenosomatidae) from Okinawa Island, Japan. Bulletin of the National Museum of Nature and Science Ser. A (Zoology), Supplement 3:175–181.
Nelson, J.S., Grande, T.C., and Wilson, M.V.H. 2016. Fishes of the World. Fifth edition. John Wiley & Sons, Hoboken.
https://doi.org/10.1002/9781119174844
Ng, P.K.L., Guinot, D., and Davie, P.J.F. 2008. Systema Brachyurorum: Part I. An annotated checklist of extant brachyuran crabs of the world. The Raffles Bulletin of Zoology, Supplement No. 17:1–286.
Ossó, À. and Gagnaison, C. 2019. An appraisal of the Middle-Late Miocene fossil decapod crustaceans of the ‘Fa-luns’ (Anjou-Touraine, France). Geodiversitas, 41(9):367–383.
https://doi.org/10.5252/geodiversitas2019v41a9
Petrascheck, W. 1952. Der Einfluß der Fazies der Flözablagerung auf die Eigenschaften der Kohle. Zeitschrift der deutschen geologischen Gesellschaft, 104:1–9.
https://doi.org/10.1127/zdgg/104/1952/1
Placer, L. 1999. Structural meaning of the Sava folds. Geologija, 41:191–221.
https://doi.org/10.5474/geologija.1998.012
Poore, G.C.B., Guinot, D., Komai, T., and Naruse, T. 2016. Reappraisal of species attributed to Halicarcinus White, 1846 (Crustacea: Decapoda: Brachyura: Hymenosomatidae) with diagnosis of four new genera and one new species from New Ireland, Papua New Guinea. Zootaxa, 4093:480–514.
https://doi.org/10.11646/zootaxa.4093.4.2
Rathbun, M.J. 1909. New crabs from the Gulf of Siam. Proceedings of the Biological Society of Washington, 22:107–114.
Schweitzer, C.E., Iturralde-Vinent, M., Hetler, J.H., and Vélez-Juarbe, J. 2006. Oligocene and Miocene decapods (Thalassinidea and Brachyura) from the Caribbean. Annals of Carnegie Museum, 75(2):111–136.
https://doi.org/10.2992/0097-4463(2006)75[111:OAMDTA]2.0.CO;2
Takeda, M. and Miyake, S. 1971. Two new hymenosomatid crabs of the genus Rhynchoplax from the West and South Pacific. Researches on Crustacea, 4:164–172.
White, A. 1846. Notes on four new genera of Crustacea. Annals and Magazine of Natural History (ser. 1), 18:176–178.
