

Article Search
Volume 27.2
May–August 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.2 May-August 2024
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
Matthew C. Mihlbachler Department of Anatomy
Department of Anatomy
New York College of Osteopathic Medicine at the New York Institute of Technology
Old Westbury, New York 11568
USA
Matthew Mihlbachler’s primary career interest is the evolutionary paleobiology of Cenozoic mammals. He has conducted field research in Asia, Africa, and throughout North America. His research is focused on the systematics, dietary paleoecology, biogeography, functional morphology, and paleopolulation demography of perissodactyls, with particular interest in understanding the role of climate change in these aspects of evolutionary biology. However, his interests in climate change and evolution has led him into investigations of other groups such as proboscideans, rodents, artiodactyls, and early condylarths. Presently, Mihlbachler is focused on understanding dental wear as a proxy for animal-environment interaction to test hypotheses about climate and morphological adaptations. He is also currently involved in field activities in the late Eocene-Oligocene transition in Mongolia and northwestern North America to better understand intercontinental faunal dynamics during that time interval.
Brian L. Beatty Department of Anatomy
Department of Anatomy
New York College of Osteopathic Medicine at the New York Institute of Technology
Old Westbury, New York 11568
USA\">
Brian Beatty's research is focused on exploring paleobiological methods, especially on secondarily aquatic tetrapods as model systems to explore the nature of functional morphology and convergence. He is especially interested in variation and the physical interactions of animals with their environment as manifest in their feeding ecology, tooth function, enamel microstructure, dental microwear, and oral pathology. Work refining paleobiological methods has led to a focus on modern aquatic groups (including Sirenia, Cetacea, Pinnipedia, Lutrinae, Crocodylia, and Chelonia) as well as terrestrial groups (Artiodactyla, Perissodactyla, Rodentia, Squamata). Fossil groups in which he is exploring how these methods can be applied include such disparate groups as Desmostylia, Embrithopoda, Multituberculata, Phytosauria, Metriorhynchoidea, Placodontia, and Mosasauria.
Angela Caldera-Siu Department of Anatomy
Department of Anatomy
New York College of Osteopathic Medicine at the New York Institute of Technology
Old Westbury, New York 11568
USA">
Angela Caldera-Siu is a medical student at the New York College of Osteopathic Medicine and joined the Department of Anatomy microwear team in 2010. She received her B.S. from Tufts University. Her past research investigated obesity and dopaminergic pathways. She plans to specialize in internal medicine.
Doris Chan Department of Anatomy
Department of Anatomy
New York College of Osteopathic Medicine at the New York Institute of Technology
Old Westbury, New York 111566
USA">
Doris Chan is currently a third year medical student at New York College of Osteopathic Medicine (NYCOM). She has conducted previous research in fields of biology and psychology. Such experiences have lead her from dissecting and analyzing mosquito midguts to recruiting and analyzing artists vs. non-artists. She joined the Angiographic Core Laboratory team at the Cardiovascular Research Foundation and studied the efficacy of drug eluting stents. In addition to her academic duties, she also runs an online website selling handmade handbags with her childhood best friend and hopes to continue doing far into the future of her medical career.
Richard Lee Department of Anatomy
Department of Anatomy
New York College of Osteopathic Medicine at the New York Institute of Technology
Old Westbury, New York 11568
USA
Richard Lee is a third year medical student at New York College of Osteopathic Medicine (NYCOM). He has participated in prior research involving Fetal Alcohol Syndrome at Binghamton University. Prior research experience involved, dissecting and monitoring of rodent models. He met Dr. Milhbachler and Dr. Beatty during his Human Anatomy course at which point he showed an interest in microwear. He worked closely with them during the second half of his first academic year. In addition to his research duties he is also a former NYCOM chapter Chief financial Officer and current member of the Student National Medical Association.
TABLE 1. P values for ANOVAs for intraobserver results from iterations 1, 2, and 3. Bold results indicate a significant difference for the same observer between iterations.
Abbreviations: NS = narrow scratches, SP = small pits, WS = wide scratches, LP = large pits.
|
|
Obs 1 |
Obs 2 |
Obs 3 |
Obs 4 |
Obs 5 |
|
NS |
0.51 |
0.30 |
0.00 |
0.00 |
0.16 |
|
SP |
0.48 |
0.12 |
0.00 |
0.16 |
0.70 |
|
WS |
0.95 |
0.05 |
0.00 |
0.01 |
0.37 |
|
LP |
0.76 |
0.98 |
0.06 |
0.03 |
0.06 |
|
WS+LP |
0.91 |
0.10 |
0.00 |
0.00 |
0.14 |
TABLE 2.P values for Least Significant Difference (LSD) and Games and Howell (GH) posthoc tests of the intraobserver data comparing adjacent iterations. Significant results are in bold. Observer-variable combinations are not shown when none of the post hoc tests were significant. LSD = least significant difference test; GH = Games and Howell test.
|
|
Iteration |
1 vs. 2 |
2 vs. 3 |
||
|
|
|
LSD |
GH |
LSD |
GH |
|
Obs 1 |
NS |
0.28 |
0.53 |
0.37 |
0.67 |
|
Obs 1 |
SP |
0.36 |
0.68 |
0.84 |
0.98 |
|
Obs 1 |
WS |
0.75 |
0.68 |
0.81 |
0.98 |
|
Obs 1 |
LP |
1.00 |
1.00 |
0.53 |
0.81 |
|
Obs 1 |
WS+LP |
0.76 |
0.95 |
0.91 |
0.99 |
|
Obs 2 |
NS |
0.79 |
0.94 |
0.15 |
0.41 |
|
Obs 2 |
SP |
0.08 |
0.08 |
0.92 |
1.00 |
|
Obs 2 |
WS |
0.04 |
0.09 |
0.89 |
0.06 |
|
Obs 2 |
LP |
0.93 |
1.00 |
0.93 |
1.00 |
|
Obs 2 |
WS+LP |
0.07 |
0.15 |
0.95 |
1.00 |
|
Obs 3 |
NS |
0.29 |
0.57 |
0.02 |
0.05 |
|
Obs 3 |
SP |
0.91 |
0.92 |
0.00 |
0.01 |
|
Obs 3 |
WS |
0.57 |
0.81 |
0.00 |
0.00 |
|
Obs 3 |
LP |
0.61 |
0.62 |
0.07 |
0.28 |
|
Obs 3 |
WS+LP |
0.45 |
0.65 |
0.00 |
0.01 |
|
Obs 4 |
NS |
0.82 |
0.84 |
0.00 |
0.03 |
|
Obs 4 |
SP |
0.95 |
0.99 |
0.09 |
0.31 |
|
Obs 4 |
WS |
0.01 |
0.01 |
0.01 |
0.02 |
|
Obs 4 |
LP |
0.01 |
0.04 |
0.51 |
0.68 |
|
Obs 4 |
WS+LP |
0.00 |
0.00 |
0.01 |
0.02 |
|
Obs 5 |
NS |
0.07 |
0.16 |
0.14 |
0.25 |
|
Obs 5 |
SP |
0.49 |
0.80 |
0.94 |
1.00 |
|
Obs 5 |
WS |
0.55 |
0.84 |
0.16 |
0.33 |
|
Obs 5 |
LP |
0.56 |
0.86 |
0.08 |
0.28 |
|
Obs 5 |
WS+LP |
0.67 |
0.92 |
0.06 |
0.14 |
TABLE 3.P values for paired t-tests for intraobserver results from iterations 1 and 2 and for iterations 2 and 3. Bold results indicate a significant difference for the same observer between iterations.
|
Iteration 1 vs. 2 |
Obs 1 |
Obs 2 |
Obs 3 |
Obs 4 |
Obs 5 |
|
NS |
0.10 |
0.50 |
0.17 |
0.48 |
0.02 |
|
SP |
0.36 |
0.03 |
0.73 |
0.91 |
0.38 |
|
WS |
0.57 |
0.00 |
0.38 |
0.00 |
0.24 |
|
LP |
1.00 |
0.91 |
0.09 |
0.01 |
0.56 |
|
WS+LP |
0.65 |
0.04 |
0.26 |
0.00 |
0.51 |
|
Iteration 2 vs. 3 |
|
|
|
|
|
|
NS |
0.05 |
0.01 |
0.02 |
0.00 |
0.00 |
|
SP |
0.81 |
0.85 |
0.00 |
0.14 |
0.92 |
|
WS |
0.48 |
0.63 |
0.00 |
0.00 |
0.00 |
|
LP |
0.13 |
0.83 |
0.04 |
0.25 |
0.03 |
|
WS+LP |
0.72 |
0.76 |
0.00 |
0.00 |
0.00 |
TABLE 4. P values for Pearson correlation coefficients comparing intraobserver results from iterations 1 and 2 and for iterations 2 and 3. Bold values indicate a significant correlation (P≤0.05).
|
Iteration 1 vs. 2 |
Obs 1 |
Obs 2 |
Obs 3 |
Obs 4 |
Obs 5 |
|
NS |
0.67 |
0.78 |
0.53 |
0.40 |
0.64 |
|
SP |
0.54 |
0.62 |
0.23 |
0.25 |
0.57 |
|
WS |
0.69 |
0.71 |
0.61 |
0.56 |
0.79 |
|
LP |
0.88 |
0.39 |
0.78 |
0.60 |
0.42 |
|
WS+LP |
0.56 |
0.39 |
0.52 |
0.62 |
0.65 |
|
Iteration 2 vs. 3 |
|
|
|
|
|
|
NS |
0.86 |
0.90 |
0.21 |
0.88 |
0.90 |
|
SP |
0.65 |
0.87 |
0.73 |
0.14 |
0.24 |
|
WS |
0.89 |
0.93 |
0.84 |
0.62 |
0.95 |
|
LP |
0.86 |
0.80 |
0.92 |
0.68 |
0.76 |
|
WS+LP |
0.91 |
0.70 |
0.81 |
0.55 |
0.94 |
TABLE 5.The number of significant (P≤0.05) Least Significant Difference (LSD) and Games and Howell (GH) post hoc tests for ANOVAs performed on the interobserver data. For each cell there are 10 possible pairwise interobserver comparisons and a total of 50 possible comparisons per iteration. The percent significant comparisons out of all possible comparisons are given in parentheses.
|
|
Iteration 1 |
Iteration 2 |
Iteration 3 |
|||
|
|
I1 LSD |
I1 GH |
I2 LSD |
I2 GH |
I3 LSD |
I3 GH |
|
NS |
6 (60%) |
7 (70%) |
4 (40%) |
3 (30%) |
7 (70%) |
6 (60%) |
|
SP |
8 (80%) |
8 (80%) |
8 (80%) |
8 (80%) |
6 (60%) |
4 (40%) |
|
WS |
7 (70%) |
6 (60%) |
4 (40%) |
2 (20%) |
9 (90%) |
8 (80%) |
|
LP |
7 (70%) |
3 (30%) |
6 (60%) |
4 (40%) |
8 (80%) |
5 (50% |
|
WS+LP |
4 (40%) |
4 (40%) |
7 (70%) |
5 (50%) |
10 (100%) |
7 (70%) |
|
total |
32 (64%) |
28 (56%) |
29 (58%) |
22 (44%) |
40 (80%) |
30 (60%) |
TABLE 6. P values for paired t-tests comparing observers 2-5 to observer 1 for iteration 3. Significant tests are in bold.
|
|
Obs 1 vs. 2 |
Obs 1 vs. 3 |
Obs 1 vs. 4 |
Obs 1 vs. 5 |
|
NS |
0.35 |
0.00 |
0.00 |
0.00 |
|
SP |
0.09 |
0.00 |
0.00 |
0.00 |
|
WS |
0.00 |
0.00 |
0.00 |
0.54 |
|
LP |
0.00 |
0.14 |
0.00 |
0.00 |
|
WS+LP |
0.00 |
0.00 |
0.00 |
0.00 |
TABLE 7.The number of significant pairwise interobserver comparisons using the Pearson correlation coefficient for iterations 1-3. For each cell there are 10 possible comparisons and a total of 50 possible comparisons per iteration. The percent of significant comparisons out of all possible comparisons are given in parentheses.
|
|
Iteration 1 |
Iteration 2 |
Iteration 3 |
|
NS |
4 (40%) |
10 (100%) |
10 (100%) |
|
SP |
4 (40%) |
10 (100%) |
10 (100%) |
|
WS |
7 (70%) |
10 (100%) |
9 (90%) |
|
LP |
2 (20%) |
10 (100%) |
10 (100%) |
|
WS+LP |
0 (0%) |
10 (100%) |
10 (100%) |
|
total |
17 (34%) |
50 (100%) |
49 (98%) |
TABLE 8. P values for Pearson correlation coefficients, comparing observer 1 to observers 2-5. Significant tests are in bold.
|
Iteration 1 |
Obs 1 vs. 2 |
Obs 1 vs. 3 |
Obs 1 vs. 4 |
Obs 1 vs. 5 |
|
NS |
0.26 |
0.69 |
0.45 |
0.58 |
|
SP |
0.46 |
0.24 |
0.04 |
0.08 |
|
WS |
0.66 |
0.52 |
0.64 |
0.54 |
|
LP |
0.13 |
0.60 |
0.26 |
0.26 |
|
WS+LP |
0.15 |
0.41 |
0.41 |
0.09 |
|
average |
0.33 |
0.49 |
0.36 |
0.31 |
|
Iteration 2 |
|
|
|
|
|
NS |
0.77 |
0.64 |
0.66 |
0.62 |
|
SP |
0.77 |
0.31 |
0.54 |
0.47 |
|
WS |
0.87 |
0.73 |
0.76 |
0.65 |
|
LP |
0.84 |
0.70 |
0.44 |
0.80 |
|
WS+LP |
0.86 |
0.61 |
0.59 |
0.72 |
|
average |
0.82 |
0.60 |
0.60 |
0.65 |
|
Iteration 3 |
|
|
|
|
|
NS |
0.72 |
0.66 |
0.42 |
0.68 |
|
SP |
0.77 |
0.57 |
0.55 |
0.85 |
|
WS |
0.68 |
0.69 |
0.61 |
0.26 |
|
LP |
0.84 |
0.51 |
0.85 |
0.50 |
|
WS+LP |
0.67 |
0.29 |
0.65 |
0.69 |
|
average |
0.74 |
0.54 |
0.62 |
0.60 |
TABLE 9. Taxon-specific P values for ANOVAs and paired t-tests, and Pearson correlation coefficients (PCC) for iteration 4, comparing observers 1 and 2. Significant results are in bold.
|
|
ANOVA |
Paired t |
PCC |
|
Equus b. |
|
|
|
|
NS |
0.18 |
0.06 |
0.58 |
|
SP |
0.87 |
0.66 |
0.89 |
|
WS |
0.10 |
0.04 |
0.51 |
|
LP |
0.01 |
0.00 |
0.41 |
|
WS+LP |
0.58 |
0.47 |
0.44 |
|
Giraffa c. |
|
|
|
|
NS |
0.54 |
0.36 |
0.58 |
|
SP |
0.65 |
0.42 |
0.71 |
|
WS |
0.55 |
0.24 |
0.77 |
|
LP |
0.31 |
0.03 |
0.86 |
|
WS+LP |
0.44 |
0.13 |
0.80 |
|
Tapirus t. |
|
|
|
|
NS |
0.97 |
0.94 |
0.78 |
|
SP |
0.70 |
0.42 |
0.78 |
|
WS |
0.80 |
0.35 |
0.93 |
|
LP |
0.26 |
0.01 |
0.90 |
|
WS+LP |
0.27 |
0.01 |
0.93 |
|
Tapirus v. |
|
|
|
|
NS |
0.02 |
0.01 |
0.48 |
|
SP |
0.00 |
0.00 |
0.28 |
|
WS |
0.46 |
0.22 |
0.67 |
|
LP |
0.31 |
0.00 |
0.92 |
|
WS+LP |
0.50 |
0.09 |
0.85 |
|
Tayassu p. |
|
|
|
|
NS |
0.32 |
0.24 |
0.31 |
|
SP |
0.22 |
0.03 |
0.76 |
|
WS |
0.34 |
0.20 |
0.52 |
|
LP |
0.25 |
0.02 |
0.82 |
|
WS+LP |
0.59 |
0.30 |
0.76 |
|
all data |
|
|
|
|
NS |
0.58 |
0.35 |
0.66 |
|
SP |
0.08 |
0.00 |
0.71 |
|
WS |
0.19 |
0.01 |
0.77 |
|
LP |
0.04 |
0.00 |
0.91 |
|
WS+LP |
0.22 |
0.01 |
0.82 |
TABLE 10.Calculation of mean absolute percentage differences (MAPD) for five observers, iterations one through three.
|
|
Average # microwear features recognized |
Mean absolute difference |
||||||||||
|
Iteration 1 |
Obs 1 |
Obs 2 |
Obs 3 |
Obs 4 |
Obs 5 |
Mean |
Obs 1 |
Obs 2 |
Obs 3 |
Obs 4 |
Obs 5 |
Mean |
|
NS |
9.5 |
8.2 |
4.2 |
6.1 |
17.2 |
9.0 |
5.1 |
9.3 |
53.5 |
32.5 |
90.3 |
38.1 |
|
SP |
9.7 |
21.4 |
0.7 |
2.0 |
5.1 |
7.8 |
24.7 |
175.1 |
91.0 |
74.3 |
34.5 |
79.9 |
|
WS |
11.7 |
7.1 |
13.9 |
21.8 |
7.4 |
12.4 |
5.5 |
42.7 |
12.3 |
76.1 |
40.2 |
35.4 |
|
LP |
3.4 |
5.9 |
0.9 |
6.3 |
3.1 |
3.9 |
13.3 |
50.5 |
77.0 |
60.7 |
20.9 |
44.5 |
|
WS+LP |
15.2 |
13.0 |
17.8 |
28.1 |
10.6 |
16.9 |
10.3 |
23.3 |
5.1 |
65.9 |
37.4 |
28.4 |
|
Iteration 2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
NS |
14.0 |
9.0 |
7.0 |
10.0 |
11.0 |
10.2 |
37.3 |
11.8 |
31.4 |
2.0 |
7.8 |
18.0 |
|
SP |
13.0 |
12.0 |
1.0 |
1.0 |
6.0 |
6.6 |
97.0 |
81.8 |
84.9 |
84.9 |
9.1 |
71.5 |
|
WS |
13.0 |
15.0 |
15.0 |
11.0 |
11.0 |
13.0 |
0.0 |
15.4 |
15.4 |
15.4 |
15.4 |
12.3 |
|
LP |
6.0 |
8.0 |
3.0 |
2.0 |
4.0 |
4.6 |
30.4 |
73.9 |
34.8 |
56.5 |
13.0 |
41.7 |
|
WS+LP |
19.0 |
23.0 |
18.0 |
13.0 |
15.0 |
17.6 |
8.0 |
30.7 |
2.3 |
26.1 |
14.8 |
16.4 |
|
Iteration 3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
NS |
11.5 |
10.8 |
8.6 |
15.8 |
15.2 |
12.4 |
7.1 |
12.8 |
30.5 |
27.6 |
22.78 |
20.2 |
|
SP |
15.8 |
13.9 |
6.0 |
10.4 |
9.3 |
11.1 |
42.6 |
25.5 |
45.9 |
6.1 |
16.06 |
27.2 |
|
WS |
10.5 |
14.6 |
25.9 |
17.8 |
10.0 |
15.8 |
33.4 |
7.4 |
64.3 |
12.9 |
36.55 |
30.9 |
|
LP |
9.0 |
12.8 |
7.5 |
5.4 |
2.7 |
7.5 |
20.3 |
71.1 |
0.3 |
27.8 |
63.90 |
36.7 |
|
WS+LP |
19.5 |
27.4 |
33.4 |
23.2 |
11.8 |
23.1 |
15.4 |
18.8 |
44.8 |
0.6 |
48.83 |
25.7 |
TABLE 11. Calculation of mean absolute percentage differences (MAPD) for observers one and two, iterations one through four.
|
|
Average # microwear features recognized |
Mean absolute difference |
||||
|
Iteration 1 |
Obs 1 |
Obs 2 |
mean |
Obs 1 |
Obs 2 |
mean |
|
NS |
9.50 |
8.20 |
8.85 |
7.34 |
7.34 |
7.34 |
|
SP |
9.70 |
21.40 |
15.55 |
37.62 |
37.62 |
37.62 |
|
WS |
11.70 |
7.10 |
9.40 |
24.47 |
24.47 |
24.47 |
|
LP |
3.40 |
5.90 |
4.65 |
26.88 |
26.88 |
26.88 |
|
WS+LP |
15.20 |
13.00 |
14.10 |
7.80 |
7.80 |
7.80 |
|
Iteration 2 |
|
|
|
|
|
|
|
NS |
14.00 |
9.00 |
11.50 |
21.74 |
21.74 |
21.74 |
|
SP |
13.00 |
12.00 |
12.50 |
4.00 |
4.00 |
4.00 |
|
WS |
13.00 |
15.00 |
14.00 |
7.14 |
7.14 |
7.14 |
|
LP |
6.00 |
8.00 |
7.00 |
14.29 |
14.29 |
14.29 |
|
WS+LP |
19.00 |
23.00 |
21.00 |
9.52 |
9.52 |
9.52 |
|
Iteration 3 |
|
|
|
|
|
|
|
NS |
11.50 |
10.80 |
11.15 |
3.14 |
3.14 |
3.14 |
|
SP |
15.80 |
13.90 |
14.85 |
6.40 |
6.40 |
6.40 |
|
WS |
10.50 |
14.60 |
12.55 |
16.33 |
16.33 |
16.33 |
|
LP |
9.00 |
12.80 |
10.90 |
17.43 |
17.43 |
17.43 |
|
WS+LP |
19.50 |
27.40 |
23.45 |
16.84 |
16.84 |
16.84 |
|
Iteration 4 |
|
|
|
|
|
|
|
NS |
11.30 |
10.60 |
10.95 |
3.20 |
3.20 |
3.20 |
|
SP |
21.30 |
17.00 |
19.15 |
11.23 |
11.23 |
11.23 |
|
WS |
14.90 |
13.00 |
13.95 |
6.81 |
6.81 |
6.81 |
|
LP |
12.60 |
17.40 |
15.00 |
16.00 |
16.00 |
16.00 |
|
WS+LP |
27.50 |
30.40 |
28.95 |
5.01 |
5.01 |
5.01 |
FIGURE 1. The "trophic triangle" of ungulate dental microwear that results from plotting scratch and pit frequencies in bivariate space, based on data from Solounias and Semprebon (2002). Each data point is the average for a species sample. Excepting a few outliers, the shaded regions represent the areas occupied by the main clusters of the three dietary categories, grazer, browser, and frugivore/hard object feeder.
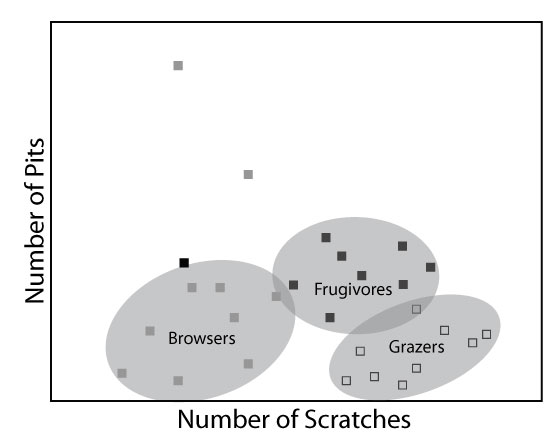
FIGURE 2. The microscope and camera used in this study, demonstrating optical and digital magnification. Digital magnification is expressed in terms of the number of pixels per micron of tooth surface. The resulting pixel width is 0.74 μm. Note that the camera does not see through the 10x eyepiece and therefore has a lower optical magnification, but nonetheless produces a resolution similar to the specimen seen through the eyepiece at 100x
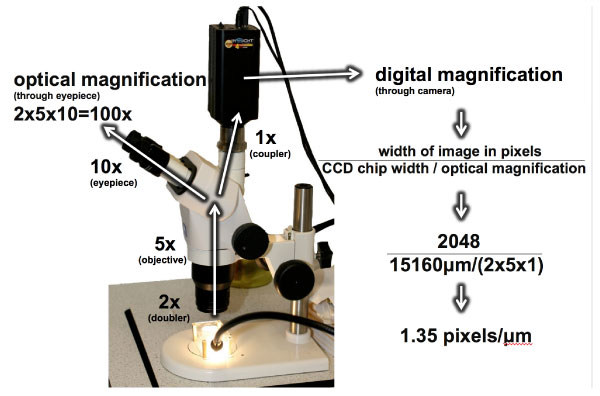
FIGURE 3. Images of two dental wear surfaces and different interpretations of three observers (Obs1, Obs 2, and Obs 3). The diameters and widths of the circles and lines with which microwear features have been marked indicate the size and shape category to which each microwear feature was assigned.

FIGURE 4. Plots of species means and standard errors for all observers during iteration 3. Browsing species are gray, grazers are white, and the frugivore/hard object feeder is black. Dotted lines represent the shape of the "trophic triangle" of each observer relating the spatial relationships of these dietary categories in bivariate space. G = giraffe (Giraffa camelopardalis), H = hippo (Hippopotamus amphibius), M = moose (Alces alces), P = peccary (Tayassu pecari), R = black rhino (Diceros bicornis), Z = zebra (Equus burchelli).
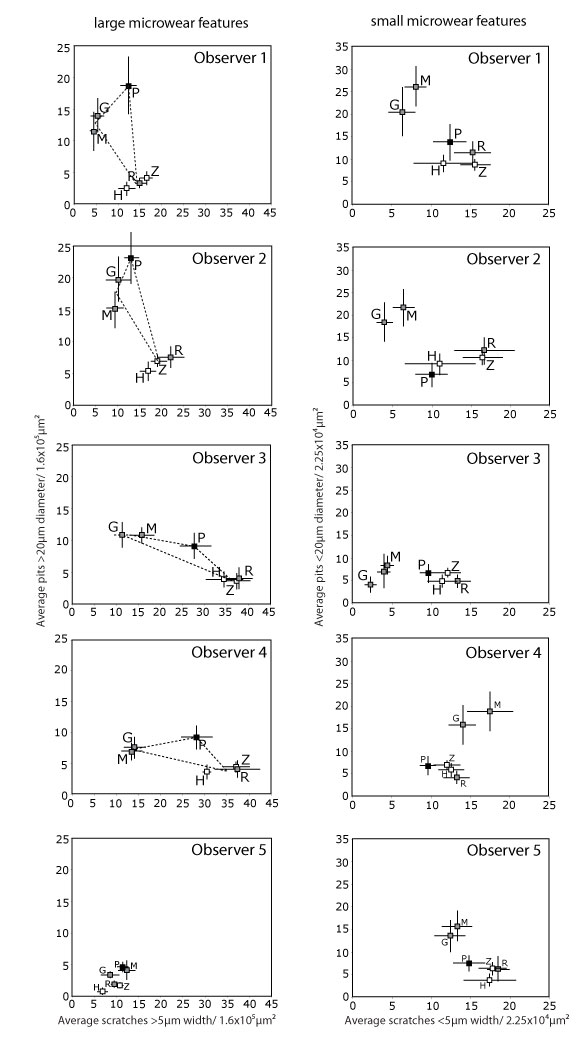
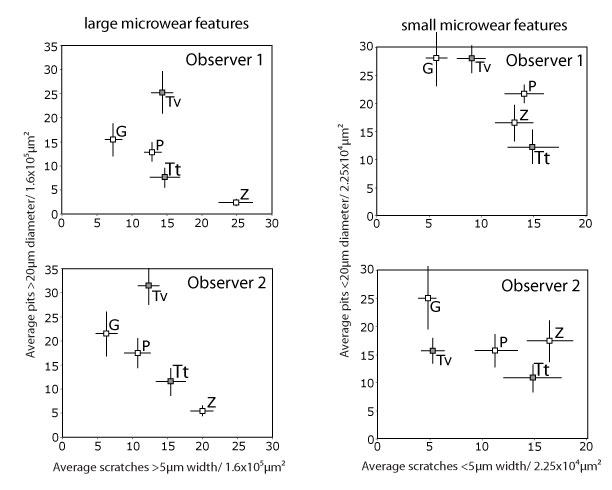
FIGURE 5. Plots of species means and standard errors for observers 1 and 2 during the fourth and final iteration. The scatter pattern for large microwear features is very similar between observers, while the scatter pattern for small microwear features is quite different. The two tapir species are gray, and other species are white. Abbreviations: G = giraffe (Giraffa camelopardalis), P = peccary (Tayassu pecari), TT = extant Brazilian tapir (Tapirus terrestris), TV = extinct Florida tapir (Tapirus veroensis), Z = zebra (Equus burchelli).
A revision of the Devonian Malvinokaffric dalmanitid trilobite Dalmanitoides Delo, 1935, on the basis of new data from Argentina
Paleontologists often interpret the past diets of extinct species by analyzing dental microwear, the microscopic wear features on fossil teeth. To analyze dental microwear, observers view wear surfaces either through the microscope or on images taken through a microscope and categorize microwear features as pits and scratches. However, microwear analysis is a subjective process, and despite significant potential for variation in how different observers perceive microwear, there has been little investigation on the magnitude of observer bias. In this study, five observers with variable amounts of initial experience with dental microwear analysis analyzed standardized sets of dental microwear images, and their results were compared iteratively as they accumulated additional experience and training. The results suggest that different observers interpret microwear patterns differently, regardless of the degree of experience or amount of practice. However, after a small amount of initial practice, all observers got very highly correlated results. In other words, even though observers did not recognize the same numbers of pits and scratches on each image, they were able to find the same relative differences between each image. These results suggest that the number of pits and scratches is not an effective means for interpreting dental microwear and is likely to lead to erroneous dietary interpretations. However, when one observer collects all the data and focuses on relative differences between microwear patterns rather than absolute numbers of microwear features, the interpretations are much more reliable.
Resumen en Español
Tasas de error y sesgo del observador en el análisis del microdesgaste dentario mediante microscopía óptica
A pesar del uso generalizado de la microscopía óptica en el análisis de microdesgaste dentario, son raros los estudios en los que se tiene en cuenta el error del observador. Sin embargo, el análisis del microdesgaste mediante microscopía óptica puede ser más propenso al sesgo del observador que el realizado mediante microscopía electrónica de barrido (SEM) o microscopía confocal. En este trabajo hemos medido el error del observador entre cinco observadores que analizaron independientemente series de superficies de desgaste dentario en microfotografías digitales tomadas mediante un microscopio óptico. En el caso de observadores experimentados, el error en el microdesgaste medido a partir del microscopio óptico es de magnitud similar a las tasas de error en los análisis mediante SEM. Un error intraobservador significativo es raro entre los observadores más experimentados y más frecuente entre los observadores no experimentados. Sin embargo, ya que los observadores se familiarizan con una imagen cuando la analizan repetidamente, el error intraobesrvador medido en este caso y en otros estudios similares mediante SEM puede ser artificialmente bajo. El error interobservador es altamente significativo y no parece que disminuya con la experiencia. No obstante, los datos recogidos por todos los observadores son estrechamente correlacionables. En lo esencial, todos los observadores encontraron diferencias similares en el microdesgaste entre las especies analizadas, aunque los valores absolutos de los datos dependen del observador. Con vistas al futuro, los resultados sobre microdesgaste serían más consistentes si los observadores adoptaran métodos que aseguraran la “ceguera” del observador y evitaran la práctica común de combinar datos de observadores diferentes, o incluso del mismo observador cuando las observaciones están separadas en el tiempo o con un grado diferente de experiencia. En lugar de utilizar bases de datos sobre microdesgaste previamente publicadas de una especie viva para interpretar paleodietas, sería más conveniente disponer de bibliotecas compartidas de imágenes de microdesgaste a partir de las cuales los observadores podrían generar sus propios datos como base de inferencias sobre las paleodietas.
PALABRAS CLAVE: desgaste dentario; microscopía óptica; microdesgaste; sesgo del observador; error del observador; paleodieta
Traducción: Miguel Company
Résumé en Français
Taux d’erreurs et biais de l’observateur dans les analyses de micro usure dentaire utilisant la microscopie optique
Malgré l’usage croissant de la microscopie optique dans les analyses de micro usure, des études montrent que les erreurs dues à l’observateur sont rares. Malgré tout, les analyses de micro usure en microscopie optique pourraient plus être sujet à des biais de l’observateur qu’avec la microscopie électronique ou confocale. Nous avons mesuré les erreurs d’observation de cinq observateurs, qui on analysé indépendamment un jeu identique de surfaces d’usure dentaire à partir de micrographies numériques prises sur un microscope optique. Pour des utilisateurs de microscopie expérimentés, les taux d’erreurs en microscopie optique furent du même ordre de grandeur que des taux d’erreurs par des méthodes utilisant la microscopie électronique. Les erreurs significatives pour un même observateur étaient rares pour les plus expérimentés et plus importantes parmi les observateurs inexpérimentés. Toutefois, du fait que les observateurs développent des habitudes liées à l’analyse répétitive de la même image, les erreurs mesurées ici pour un même observateur et dans des études similaires en microscopie électronique peuvent être artificiellement basses. Les erreurs entre observateurs étaient hautement significatives et ne semblaient pas diminuer avec l’expérience. Toutefois les données collectées par les différents observateurs étaient hautement corrélées. De façon générale tous les observateurs on trouvé des différences de micro usure équivalentes entre les espèces analysées, bien que les valeurs absolues de données étaient dépendantes des observateurs. A l’avenir, les résultats de micro usure seront plus robustes si les observateurs adoptent des méthodes qui assurent l’objectivité de l’observateur, et évitent la pratique courante de combiner les données provenant de différents observateurs, ou même des données d’un même observateur quand ses observations sont séparées par le temps et l’expérience. Plutôt que d’utiliser des bases de données pré-publiées de micro usure dentaire d’espèces actuelles comme référence pour interpréter les habitudes alimentaires passées, les chercheurs auraient meilleur compte de construire une librairie commune d’images de micro usure à partir de laquelle les observateurs pourraient générer leur propre données de base pour interpréter les habitudes alimentaires dans le fossile.
MOTS CLEFS : usure dentaire, microscopie optique, micro usure dentaire, biais de l’observateur, alimentation passée.
Translator: Olivier Maridet
Deutsche Zusammenfassung
Fehlerquoten und Betrachterverzerrung bei Microwear–Analysen mit Lichtmikroskopie
Trotz des gestiegenen Einsatzes von Lichtmikroskopie bei Mikrowear-Analysen sind Untersuchungen über Betrachterfehler selten. Dennoch ist Mikrowear-Analyse mit Lichtmikroskopie möglicherweise anfälliger für Betrachterfehler als SEM oder Konfokalmikroskopie. Wir maßen Betrachterfehler bei fünf Betrachtern, die unabhängig voneinander identische Sätze digitaler Aufnahmen von dentalen Microwear-Oberflächen analysierten, die durch einem Lichtmikroskop gemacht worden waren. Für erfahrene Mikroskopiker war der Fehler bei der Lichtmikroskopie-basierten Microwear gleich hoch wie die Fehlerquoten bei SEM-basierten Microwear-Methoden. Signifikante Intraobserver-Fehler waren bei den erfahrensten Betrachtern selten und höher bei den Unerfahrenen. Da jedoch Betrachter bei wiederholtem analysieren desselben Bildes eine Vertrautheit entwickeln, mag die hier und bei anderen ähnlichen SEM-Untersuchungen gemessene Intraobserver-Fehlerquote künstlich niedrig sein. Interobserver-Fehler waren von hoher Relevanz und schienen sich bei mehr Erfahrung nicht zu verringern. Nichtsdestotrotz korrelierten die von allen Betrachtern gesammelten Daten eng miteinander. Im Wesentlichen fanden alle Betrachter ähnliche Microwear-Unterschiede zwischen den analysierten Arten obwohl die Absolutwerte vom jeweiligen Betrachter abhingen. In Zukunft werden Microwear-Analysen stabiler sein, wenn die Betrachter Methoden einführen, die „Betrachterblindheit“ sicherstellen. Auch sollte die übliche Praxis der Datenkombination von verschiedenen Betrachtern oder einem einzelnen Betrachter, wenn die Beobachtungen über einen längeren Zeitraum durchgeführt oder unterbrochen wurden, vermieden werden. Statt bereits publizierte Microwear-Datenbanken über heutige Arten als Basis für die Bestimmung von Paläonahrung zu nutzen, könnte es für Wissenschaftler besser sein, gemeinsam nutzbare Microwear-Bilderverzeichnisse aufzubauen mit denen Betrachter ihre eigenen Daten generieren und Schlussfolgerungen über Paläonahrung anstellen können
SCHLÜSSELWÖRTER: Zahnabrieb; Lichtmikroskopie; Microwear; Betrachterverzerrung; Betrachterfehler; Paläonahrung
Translators: Eva Gebauer and Anke Konietzka
Arabic

Translator: Ashraf M.T. Elewa
Polski Abstrakt
WSKAŹNIKI BŁĘDÓW I ODCHYLENIE OBSERWACJI BADACZA W ANALIZIE MIKROZUŻYCIA ZĘBÓW Z WYKORZYSTANIEM MIKROSKOPII ŚWIETLNEJ
Pomimo coraz większego wykorzystywania mikroskopii świetlnej w analizie mikrozużycia zębów, ilość badań skupiających się na określeniu błędów popełnianych przez osoby wykonujące te analizy jest stosunkowo niewielka. Pomimo tego analiza mikrozużycia z wykorzystaniem mikroskopii świetlnej może być bardziej podatna na subiektywizm badaczy niż SEM czy mikroskopia konfokalna. Zmierzyliśmy błędy popełniane wśród grupy pięciu badaczy, którzy niezależnie analizowali identyczne zestawy powierzchni zużycia zębów na mikrofotografiach wykonanych przy pomocy mikroskopu świetlnego. W przypadku doświadczonych badaczy pracujących z mikroskopem wielkości błędów analiz wykonanych mikroskopem świetlnym były porównywalne do błędów powstałych przy wykorzystaniu SEM. Ilość znaczących błędów poszczególnych badaczy była niższa u doświadczonych naukowców a wyższa u niedoświadczonych badaczy. Jednakże ponieważ ciągłe obserwowanie tych samych próbek powoduje zapoznanie się z nimi, zmierzone w tej i innych podobnych analizach SEM błędy poszczególnych badaczy mogą być sztucznie zaniżone. Błędy pomiędzy badaczami były wyraźnie znaczące i nie wykazywały spadku wraz ze wzrostem doświadczenia. Pomimo tego wyniki otrzymane przez wszystkich naukowców były ściśle korelowalne. Zasadniczo wszyscy badacze znaleźli te same różnice w mikrozużyciu pomiędzy badanymi gatunkami, nawet pomimo faktu iż absolutne wartości danych były zależne od konkretnego naukowca. Kontynuując, wyniki badań mikrozużycia będą bardziej dokładne jeśli badacze zaczęliby stosować metodologie, które zapewniłyby niezależność naukowca od poprzednich wyników badań, zarówno jeśli chodzi o łączenie wyników otrzymanych przez innych badaczy jak również swoich oddzielonych czasem i poziomem doświadczenia. Zamiast używać opublikowanych wcześniej wyników badań mikrozużycia zębów współczesnych gatunków jako bazy do interpretacji paleodiety, badacze przysłużyliby się lepiej tworząc ogólnodostępną bazę zdjęć mikrozużycia, dzięki którym inni naukowcy mogliby generować ich własne dane jako bazy do interpretacji paleodiety.
Słowa kluczowe: zużycia zębów, mikroskopia świetlna, mikrozużycie, odchylenie wyników badaczy, błędy badaczy, paleodieta
Translators: Dawid Mazurek, Robert Bronowicz, and Daniel Madzia
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024