

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Carolin Haug
Carolin Haug
Department of Geology and Geophysics
Yale University
Kline Geology Laboratory
210 Whitney Avenue
New Haven, Connecticut 06511
USA
Current address: Zoological Institute and Museum, Cytology and Evolutionary Biology, University of Greifswald, Soldmannstr. 23, 17487 Greifswald, Germany
Carolin Haug studied biology at the Julius-Maximilians-Universität Würzburg with animal ecology as major subject and sociobiology and palaeontology as minor subjects. She received her Diplom in 2005 and moved to the University of Ulm for her PhD thesis. In the work group Biosystematic Documentation she investigated the ontogeny and evolution of the arthropod head shield on a wide spectrum of fossil and extant specimens. Due to the very different demands of the material, she developed new imaging techniques or modified existing methods together with her collaborators, for example, composite imaging, stereo photography or autofluorescence macro- and microscopy. In 2011, Carolin defended her PhD thesis. She is currently as postdoctoral researcher at Yale University. Besides optimising imaging techniques, she is interested in fossilised development (see also http://www.palaeo-evo-devo.info) and in the evolution of tagmosis. For this purpose, she investigates mainly arthropods from different deposits yielding exceptional preservation, such as Rhynie chert, Solnhofen Lithographic Limestones, the 'Orsten', Burgess Shale, or Mazon Creek, always in comparison to their extant relatives. Carolin will move to Greifswald in June 2012 to continue her research there at the Ernst-Moritz-Arndt-Universität.
![]()
Joachim T. Haug Department of Geology and Geophysics
Department of Geology and Geophysics
Yale University
Kline Geology Laboratory
210 Whitney Avenue
New Haven, Connecticut 06511
USA
Current address: Zoological Institute and Museum, Cytology and Evolutionary Biology, University of Greifswald, Soldmannstr. 23, 17487 Greifswald, Germany
Joachim T. Haug studied animal ecology (major), sociobiology and palaeontology (minors) at the Julius-Maximilians-Universität Würzburg. After the receipt of the Diplom in 2005, he moved to the work group Biosystematic Documentation at the University of Ulm for his PhD thesis. There he worked on the early crustaceans from the Cambrian 'Orsten', which are preserved three-dimensionally and with minute details. To make the entire morphology clearly visible, Joachim produced computer-based 3D models for each developmental stage of a species, resulting in a 4D model of the species. With 4D models reconstructed for the different species, differences in the developmental pattern between the species became visible. These changes in the developmental timing, so-called heterochronic events must have occurred several times during early crustacean evolution. Due to these findings, Joachim got interested in studying such evolutionary changes of development also in fossils from other deposits and focussed on Palaeo-Evo-Devo (see also http://www.palaeo-evo-devo.info). After his defense in 2009 and two further years as postdoctoral researcher in Ulm, he received a Feodor Lynen research fellowship from the Alexander von Humboldt-Foundation, with which he is currently doing research at Yale University. Besides 3D modelling, Joachim is also interested in different methods of imaging (documentation and presentation). In June 2012, he will relocate to the Ernst-Moritz-Arndt-Universität Greifswald.
![]()
Stephen R. Fayers Department of Geology and Petroleum Geology
Department of Geology and Petroleum Geology
School of Geosciences
University of Aberdeen
Meston Building
Kings College
Aberdeen, AB24 3UE
UK
Stephen Fayers studied geology as an undergraduate in Portsmouth and went on to complete a master’s degree, gaining a distinction in Sedimentology and its Applications at the University of Reading. Following a move to Aberdeen in Scotland to work as a sedimentologist in the oil and gas industry, Stephen returned to academia in 1999 to study the sedimentology, biota and palaeoenvironments of the Windyfield chert; part of a complex of Early Devonian (Pragian-age) terrestrial hot spring deposits found at Rhynie in NE Scotland. Finishing his Ph.D. in 2003, Stephen continued his work, in collaboration with numerous international universities, with the Rhynie Research Group at the University of Aberdeen as a Post-Doctorate Research Fellow. Since this time he has written and co-authored numerous research papers in peer-reviewed journals. Stephen currently works as a Senior Geoscientist for Baker Hughes in Aberdeen, specialising in borehole image log analysis; he also teaches borehole image log processing and interpretation at M.Sc. level for the University of Aberdeen. His primary research interests continue to centre around the palaeoenvironments, biota and palaeoecology of early terrestrial and freshwater ecosystems.
![]()
Nigel H. Trewin Department of Geology and Petroleum Geology
Department of Geology and Petroleum Geology
School of Geosciences
University of Aberdeen
Meston Building
Kings College
Aberdeen, AB24 3UE
UK
Nigel H. Trewin is an Emeritus Professor of Geology at Aberdeen University, Scotland, and Honorary Curator of the Geological Collections at the university. His academic papers over 44 years are mostly on sedimentology, diagenesis, palaeontology, stratigraphy and regional geology. Current specialisations are the Old Red Sandstone of Scotland, particularly the Rhynie chert, and fish-bed faunas. Published books include ‘The Geology of Scotland’ (Ed. 4th Edn 2002), an excursion guide book to East Sutherland and Caithness (2009) and ‘Fossils Alive’ (2008).
![]()
Christopher Castellani Biosystematic Documentation
Biosystematic Documentation
University of Ulm
Helmholtzstrasse 20
D-89081 Ulm
Germany
Christopher Castellani is currently a PhD student in the workgroup of Prof. Dr. Dieter Waloszek at the University of Ulm (Germany). He studied geology and paleontology at Brest, Marseille and Dijon (France), for five years. During his Master thesis, he specialized in modern morphometric and statistical techniques and used them to characterise the morphological disparity within the taxon Carabus. In 2007, he moved to the University of Ulm (Germany) to integrate the workgroup of Prof. Dr. Dieter Waloszek as Research Assistant. He was involved in a European Program, funded by the Marie Curie Early Stage Research Training, about the “Evolution of animal body plans as inferred by developmental biology, morphology, molecular phylogeny, and palaeontology” (project MOLMORPH). During this 1-year fellowship he worked on the ontogeny and life habit of the Upper Cambrian parasitic tongue worms Pentastomida found in various ’Orsten’ deposits of Sweden. Christopher has then started his PhD thesis in 2009 and he attempts to precise the palaeo-biodiversity and palaeo-ecology of the Cambrian ’Orsten’ assemblages of the Alum Shale Formation of Sweden. Previous investigations, under the direction of Prof. Dr. Klaus J. Müller and Prof. Dr. Dieter Waloszek, of the so-called ‘Orsten’-type fossils yielded a great number of immature and adult forms of mainly meiofaunic arthropods. However, the ‘Orsten’ faunas do not only comprise arthropod organisms, re-investigation of the original residues has led to the discoveries of numerous micropaleontolgical remains (mainly fragments) of various affinities (sponges, echinoderms, Small Shelly Fossils, brachiopods, graptolites, cyanobacteria, various ‘worms’ and unknown arthropods, and other problematic fossils). Christopher aims to draw a more realistic picture of the biodiversity of the ‘Orsten’ assemblages and more particularly of the biotas that inhabited the shallow Alum Shale sea during the Furongian (Upper Cambrian). His research techniques comprise mainly scanning electron microscopy due to the small size of the fossils, but also thin sections and modern imaging techniques such as synchrotron and fluorescence microscopy. Christopher is interested in various aspects of the ‘Orsten’-type Lagerstätten (biodiversity and taphonomic processes leading their preservation in Sweden and worldwide) and in general, in the biodiversity, in term of species richness, morphological disparity and ecological strategy, of early Palaeozoic ecosystems and their evolution through time.
![]()
Dieter Waloszek Biosystematic Documentation
Biosystematic Documentation
University of Ulm
Helmholtzstrasse 20
D-89081 Ulm
Germany
Dieter Waloszek is working as a professor at the University of Ulm. He studied biology in Hamburg and finished his dissertation with a thesis on a scallop from Argentinian offshore waters. In 1982 Dieter moved to the Institute of Palaeontology in Bonn to work together with Klaus J. Müller, the discoverer of the Cambrian 3d-preserved, so-called ‘Orsten’ fossils, mainly arthropods. After his habilitation in 1991 on one of the faunal elements, the crustacean Rehbachiella kinnekullensis, and the phylogeny of Branchiopoda and Crustacea and one year in Kiel, Dieter received a professorship in Ulm in 1995. Since then, he and his workgroup continue the studies of ‘Orsten’ fossils. Major method remains the SEM, but the workgroup expanded their investigations to other localities worldwide, material from extant and other fossil sources and their methodological range for improving the documentation of this important evolutionary data source.
More recent highlight: foundation of the international C.O.R.E. group of scientists with a specific interest in ‘Orsten’ studies.
Main interests: morphology, incl. functional aspects, ontogeny, phylogeny and evolution – no taxonomic restriction in general, but due to capacity problems, focus on Arthropoda.
Output (incl. team work): more than 100 papers in international journals.
Main hobbies: family, fistball, holidays in Sweden, incl. fishing and waterskiing.
![]()
Andreas Maas Biosystematic Documentation
Biosystematic Documentation
University of Ulm
Helmholtzstrasse 20
D-89081 Ulm
Germany
Andreas Maas is a research assistant at the University of Ulm. He studied biology at the Christian Albrechts University of Kiel, Germany and graduated in 1998 with a diploma thesis on the larval development of the Antarctic krill Euphausia superba. In 2002 he made his PhD in zoology at the University of Ulm. Andreas studied a Cambrian group of crustaceans, the Phosphatocopina; their systematic position within Crustacea could be established as the sister group of the crustacean crown group, the Eucrustacea. The material of his study is part of the so-called ‘Orsten’ type of preservation that leads to three-dimensional fossils of mainly less than 1 mm in size. Since 2002 Andreas is also involved in teaching at the University of Ulm and did his habilitation in 2008. He was part of a German-Chinese research cooperation and was able to work on fossils from the famous lower Cambrian Chengjiang fauna. With this Andreas consolidated his main research subject in the early evolution and phylogeny of arthropods. He could widen this subject also on the question of the phylogenetic position of arthropods within the Bilateria. Andreas worked on very early, worm-like arthropods without sclerotised dorsal tergites that were from the same Cambrian material as the phosphatocopines before. Another important Cambrian fossil he described was a minute, Cambrian, possibly immature stage of a new species of Cycloneuralia. Shergoldana australiensis gives some clues about the evolution of larval forms within Nemathelminthes. This is especially important since arthropods are regarded as close relatives of round worms nowadays. Andreas’ research techniques comprise mainly scanning electron microscopy due to the small size of the fossils. Important other skills are computer-aided imagery and computer-aided phylogeny analyses. Andreas teaches evolutionary biology including history and background of Darwin’s important work and evolution, systematics and phylogeny of animals, Metazoa.
![]()
TABLE 1. Measurements for all specimens in micrometers with reference to appearance on certain figures. Brackets indicate uncertainties in assignment to certain size class or in measured values due to weak preservation. Specimen AUGD 12445C is mentioned, as it was documented and received a number, but later turned out as not being part of a crustacean. Numbers in first row mean: 1 = Figure; 2 = Repository number AUGD; 3 = Size class; 4 = Diameter of mandibular exopod (d1); 5 = Diameter of antennal exopod (d2); 6 = Diameter of exopod with unclear affinities; 7 = Diameter of antennal endopod (d3); 8 = Diameter of mandibular endopod; 9 = Height of antennal coxa in proximal-distal axis (h); 10 = Length of antennal coxa in median-lateral axis (l).
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
5.1 |
12449B |
III |
61 |
61 |
– |
44 |
18 |
71 |
139 |
|
5.2 |
12448A |
II |
50 |
46 |
– |
27 |
– |
61 |
96 |
|
5.3–5 |
12449A01 |
(II) |
44 |
– |
– |
– |
17 |
– |
– |
|
6.1 |
12448B |
II |
37 |
41 |
– |
27 |
16 |
– |
– |
|
6.2 |
12449C |
II |
44 |
42 |
– |
27 |
– |
51 |
94 |
|
6.3 |
12449A25 |
I |
35 |
35 |
– |
24 |
14 |
– |
– |
|
6.4 |
12449A02 |
III |
61 |
53 |
– |
35 |
– |
78 |
114 |
|
6.5(left) |
12449A03a |
II |
39 |
45 |
– |
28 |
18 |
55 |
84 |
|
6.5(right) |
12449A03b |
II |
43 |
45 |
– |
29 |
21 |
58 |
84 |
|
6.6 |
12449E |
I |
31 |
31 |
– |
18 |
– |
31 |
68 |
|
6.7–8 |
12449A04 |
III |
– |
57 |
– |
32 |
– |
64 |
131 |
|
7.1 |
12445D |
II |
43 |
51 |
– |
– |
23 |
73 |
106 |
|
7.2 |
12449A20 |
II |
– |
41 |
– |
28 |
– |
61 |
94 |
|
7.3 |
12449A18 |
I |
– |
36 |
– |
23 |
– |
51 |
64 |
|
7.4 |
12451F |
II |
45 |
45 |
– |
– |
– |
65 |
96 |
|
7.5 |
12447C |
I |
– |
32 |
– |
19 |
– |
50 |
62 |
|
7.6 |
12447A |
II |
– |
46 |
– |
29 |
– |
65 |
|
|
7.7 |
12449A22 |
II |
– |
46 |
– |
20 |
– |
– |
– |
|
8.1(bottom) |
12449A05a |
II |
– |
43 |
– |
23 |
– |
– |
– |
|
8.1(top) |
12449A05b |
II |
– |
41 |
– |
28 |
– |
64 |
110 |
|
8.2 |
12449A06 |
(II) |
41 |
– |
– |
– |
– |
– |
– |
|
8.3 |
12449A08 |
I |
– |
36 |
– |
15 |
– |
– |
– |
|
8.4 |
12449A16 |
I |
34 |
34 |
– |
– |
– |
46 |
90 |
|
8.5 |
12449A09 |
III |
– |
51 |
– |
33 |
– |
– |
– |
|
8.6 |
12449A14 |
I |
– |
32 |
– |
– |
– |
43 |
85 |
|
8.7 |
12449A19 |
II |
46 |
44 |
– |
25 |
– |
– |
108 |
|
8.8 |
12447B |
II |
– |
41 |
– |
25 |
– |
71 |
99 |
|
8.9 |
12454C |
II |
– |
45 |
– |
– |
– |
49 |
106 |
|
9.1 |
12452B |
(I) |
– |
28 |
– |
– |
18 |
– |
– |
|
9.2 |
12449A21 |
(II) |
– |
– |
47 |
– |
– |
– |
– |
|
9.3 |
12449A10 |
(II–III) |
– |
– |
53 |
– |
– |
– |
– |
|
9.4 |
12451B |
I |
– |
28 |
– |
15 |
– |
– |
– |
|
9.5 |
12447F |
? |
– |
– |
45 |
– |
– |
– |
– |
|
9.6 |
12449G |
(II) |
– |
– |
44 |
– |
– |
– |
– |
|
9.7 |
12454F |
(II–III) |
– |
– |
52 |
– |
– |
– |
– |
|
9.8 |
12442E |
(II) |
– |
– |
43 |
– |
– |
– |
– |
|
9.9 |
12454D |
? |
– |
– |
– |
– |
– |
– |
– |
|
9.10 |
12451C |
II |
– |
– |
– |
– |
– |
59 |
83 |
|
9.11 |
12449A15 |
II |
– |
– |
– |
– |
– |
42 |
106 |
|
10.1 |
12442B |
I |
– |
25 |
– |
– |
– |
– |
68 |
|
10.2 |
12445A |
(II) |
– |
– |
39 |
– |
– |
– |
– |
|
10.3 |
12447E |
? |
– |
– |
– |
– |
– |
– |
– |
|
10.4 |
12447D |
(III) |
– |
– |
60 |
– |
– |
– |
– |
|
10.5 |
12444B |
(II) |
– |
– |
44 |
– |
– |
– |
– |
|
10.6 |
12448E |
(II) |
– |
– |
39 |
– |
– |
– |
– |
|
10.7 |
12448F |
(III) |
– |
– |
61 |
– |
– |
– |
– |
|
10.8 |
12449A23 |
(I) |
– |
– |
25 |
– |
– |
– |
– |
|
10.9 |
12454G |
(I) |
– |
– |
(31) |
– |
– |
– |
– |
|
10.10 |
12454I |
(I) |
– |
– |
– |
(16) |
– |
– |
– |
|
10.11 |
12454B |
(II) |
– |
39 |
– |
– |
– |
– |
– |
|
10.12 |
12454H |
(II–III) |
– |
– |
52 |
– |
– |
– |
– |
|
10.13 |
12451D |
(III) |
– |
53 |
– |
– |
– |
– |
– |
|
11.1 |
12454E |
(I) |
– |
25 |
– |
– |
– |
– |
– |
|
11.2 |
12442D |
(II) |
– |
– |
41 |
– |
– |
– |
– |
|
11.3 |
12448G |
(II) |
– |
– |
45 |
– |
– |
– |
– |
|
11.4 |
12454A |
III |
55 |
57 |
– |
– |
– |
– |
– |
|
11.5 |
12451A |
I |
34 |
36 |
– |
22 |
13 |
49 |
78 |
|
11.6 |
12445F |
II |
– |
41 |
– |
27 |
– |
61 |
102 |
|
11.7 |
12449A17 |
II |
44 |
46 |
– |
25 |
– |
– |
– |
|
11.8 |
12449A11 |
II |
– |
43 |
– |
20 |
– |
61 |
112 |
|
11.9 |
12449A07 |
II |
47 |
45 |
– |
33 |
14 |
– |
– |
|
12.1 |
12451G |
I–II |
– |
36 |
– |
– |
– |
41 |
92 |
|
12.2 |
12449D |
(II–III) |
– |
55 |
– |
– |
– |
– |
– |
|
12.3 |
12449A13 |
(II) |
– |
39 |
– |
– |
– |
– |
– |
|
12.4 |
12448C |
(II) |
– |
51 |
– |
– |
– |
– |
– |
|
12.5 |
12449F |
I |
25 |
25 |
– |
19 |
– |
28 |
55 |
|
12.6 |
12449A12 |
(II–III) |
– |
– |
54 |
– |
– |
– |
– |
|
12.7 |
12452A |
(III) |
– |
– |
59 |
– |
– |
– |
– |
|
12.8 |
12449A24 |
I |
– |
33 |
– |
19 |
– |
– |
70 |
|
12.9 |
12451E |
(I) |
– |
– |
– |
17 |
– |
– |
– |
|
13.1 |
12442A |
(II) |
– |
49 |
– |
– |
– |
– |
– |
|
13.2 |
12444A |
(II) |
– |
– |
– |
– |
– |
65 |
99 |
|
13.3 |
12442C |
II |
45 |
41 |
– |
– |
– |
– |
– |
|
13.4 |
12448D |
I |
– |
32 |
– |
– |
– |
31 |
64 |
|
13.5 |
12445E |
(II) |
– |
– |
41 |
– |
– |
– |
– |
|
13.6 |
12448H |
III |
– |
63 |
– |
33 |
– |
80 |
133 |
|
13.7 |
12445B |
(II) |
– |
– |
49 |
– |
– |
– |
– |
|
13.8 |
12447G |
III |
– |
65 |
– |
39 |
– |
65 |
139 |
|
13.9 |
12442F |
I |
– |
35 |
– |
25 |
– |
41 |
78 |
|
not a crustacean |
12445C |
– |
– |
– |
– |
– |
– |
– |
– |
TABLE 2. Overview of the most important measurement data in µm. Lengths of entire appendages or of exopods and endopods always without terminal setae. Question marks indicate uncertainties due to preservational constraints; dashes mark unavailable data.
|
Value |
Stage I |
Stage II |
Stage III |
|
total anterior-posterior body length |
150 |
225 |
300 |
|
maximum body width (excl. appendages) |
160 |
185 |
270 |
|
maximum dorso-ventral body height |
90 |
145 |
205 |
|
antennula length |
– |
90 |
150? |
|
antenna length |
190 |
270 |
370 |
|
antennal coxa, medio-lateral extension |
75 |
105 |
135 |
|
antennal coxa, maximum proximo-distal height |
40 |
60 |
75 |
|
antennal endopod length |
60 |
80 |
110 |
|
antennal exopod length |
160 |
205 |
280 |
|
mandible length |
170? |
220 |
280 |
|
mandibular coxa, medio-lateral extension |
– |
45 |
53 |
|
mandibular coxa, maximum proximo-distal height |
– |
60? |
75? |
|
mandibular endopod length |
14? |
18? |
20? |
|
mandibular exopod length |
130 |
165 |
205 |
TABLE 3. Comparative data from the first larval stage (nauplius) of selected eucrustacean species; data source given in brackets for each species separately. I. Data of the nauplius described herein. II. Rehbachiella kinnekullensis Müller, 1983 (Branchiopoda) from the Cambrian 'Orsten' of Sweden (Walossek, 1993). III. Artemia sp. from Macau strain, north-eastern Brazil (Schrehardt, 1987). IV. Bryocamptus pygmaeus (Sars, 1862) (Copepoda, Harpacticoida) from fresh water of NW Germany (Dahms, 1987). V. Pseudotachidius sp. (Copepoda, Harpacticoida) from Weddell Sea (Dahms, 1989). VI. Eudiaptomus gracilis (Sars, 1863) (Copepoda, Calanoida) from fresh water of Bavaria, Germany (unpublished data of M. Mayer in the workgroup of D. Waloszek). VII. Monstrilla hamatapex Grygier and Ohtsuka, 1995 (Copepoda, Monstrilloida) from adjacent to Tanabe Bay, Japan (Grygier and Ohtsuka, 1995). VIII. Laura bicornuta Grygier, 1985 (Ascothoracida) from Hawaii (Grygier, 1985). IX. Hansenocaris furcifera Itô, 1989 (Facetotecta) from Tanabe Bay, Japan (Itô, 1990). X. Briarosaccus tenellus Boschma, 1970 (Cirripedia, Rhizocephala) (Walossek et al., 1996). XI. Euphausia superba (Dana, 1852) (Euphausiacea) from Antarctic waters (Fraser, 1936). XII. Macropetasma africanum (Balss, 1913) (Decapoda, Penaeoidea) from off the coast along southern Africa (Cockcroft, 1985). ? = data uncertain or unknown.
|
Value |
I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
XI |
XII |
|
total length |
150 |
160 |
350 |
80 |
235 |
135 |
50 |
440 |
280 |
320 |
650 |
250 |
|
total width |
160 |
? |
120 |
65 |
200 |
75 |
32 |
280 |
170 |
200 |
480 |
130 |
|
antennula number of elements |
6 |
? |
1 |
3 |
3 |
2 |
3 |
5 |
2 |
3 |
1 |
1 |
|
antennula length |
? |
? |
50 |
25 |
50 |
80 |
12 |
180 |
40 |
130 |
300 |
200 |
|
antenna length |
190 |
100 |
200 |
60 |
100 |
100 |
15 |
200 |
80 |
155 |
320 |
200 |
|
antennal endopod number of elements |
3 |
4 |
1 |
1 |
1 |
1 |
2 |
3 |
2 |
1 |
1 |
1 |
|
antennal endopod length |
60 |
60 |
60 |
40 |
30 |
23 |
10 |
100 |
25 |
75 |
60 |
150 |
|
antennal exopod number of annuli |
12 |
7–8 |
6–8 |
2 |
5 |
7 |
4 |
8–9 |
6 |
8 |
1 |
1 |
|
antennal exopod length |
160 |
70 |
80 |
20 |
50 |
52 |
8 |
130 |
45 |
90 |
60 |
160 |
|
mandible length |
170? |
85 |
60 |
25 |
60 |
50 |
12 |
180 |
80 |
100 |
300 |
100 |
|
mandibular endopod number of elements |
1 |
3 |
1 |
1 |
1 |
1 |
1? |
3 |
2 |
1 |
1 |
1 |
|
mandibular endopod length |
14? |
45 |
10 |
10 |
10 |
23 |
10? |
100 |
25 |
30 |
60 |
70 |
|
mandibular exopod number of elements |
7 |
7 |
1 |
2 |
2 |
5 |
1 |
7 |
5 |
6 |
1 |
1 |
|
mandibular exopod length |
130 |
50 |
10 |
20 |
25 |
24 |
3 |
110 |
40 |
55 |
60 |
70 |
APPENDIX 1.
Overview of specimens on thin section AUGD 12442.

Overview of specimens on thin section AUGD 12444.

Overview of specimens on thin section AUGD 12445.

Overview of specimens on thin section AUGD 12447.

Overview of specimens on thin section AUGD 12448.

Overview of specimens on thin section AUGD 12449, not glued side.

Overview of specimens on thin section AUGD 12449, glued side.

Overview of specimens on thin section AUGD 12451.

Overview of specimens on thin section AUGD 12452.
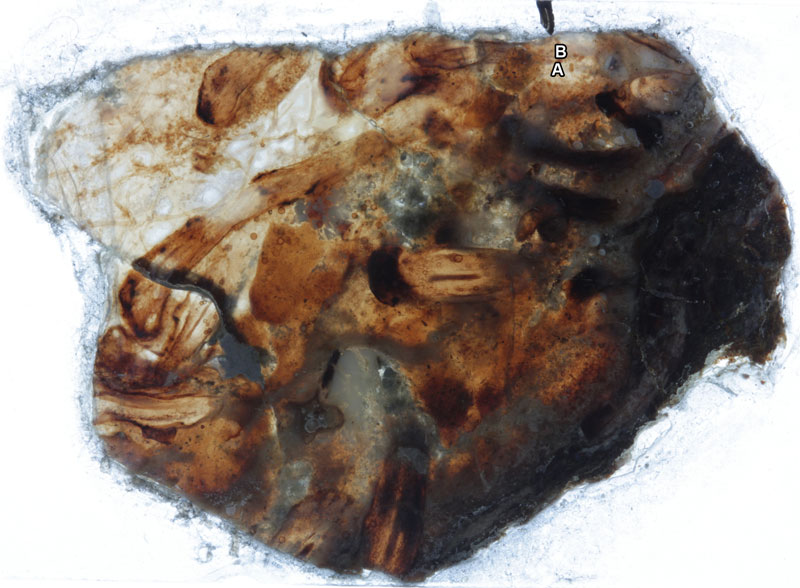
Overview of specimens on thin section AUGD 12454.

FIGURE 1. Scheme depicting the measured parts of antenna (ant) and mandible (md): d1 = diameter of the mandibular exopod (ex); d2 = diameter of the antennal exopod; d3 = diameter of the antennal endopod (en); h = height of the antennal coxa (cx) in proximal-distal axis; l = length of the antennal coxa in median-lateral axis. Other abbreviation: bas = basipod.
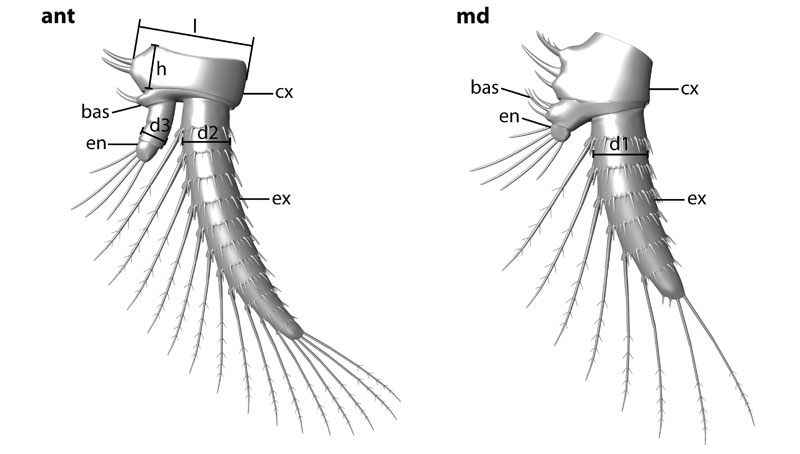
FIGURE 2. Scatter and box plots of the measured parameters. 2.1. Diameter of the mandibular exopod (d1) versus diameter of the antennal exopod (d2). 2.2. Diameter of the antennal endopod (d3) versus diameter of the antennal exopod (d2). 2.3. Height of the antennal coxa in proximal-distal axis (h) versus length of the antennal coxa in median-lateral axis (l). 2.4. Diameter of the antennal exopod (d2) versus length of the antennal coxa in median-lateral axis (l). 2.5. 3D scatter plot of specimens of which d2, d3 and l could be measured. 2.6. Box plot of parameter d2; three size classes highly significant (Wilks Lambda = 0.097, F = 12.505, p < 0.0001).
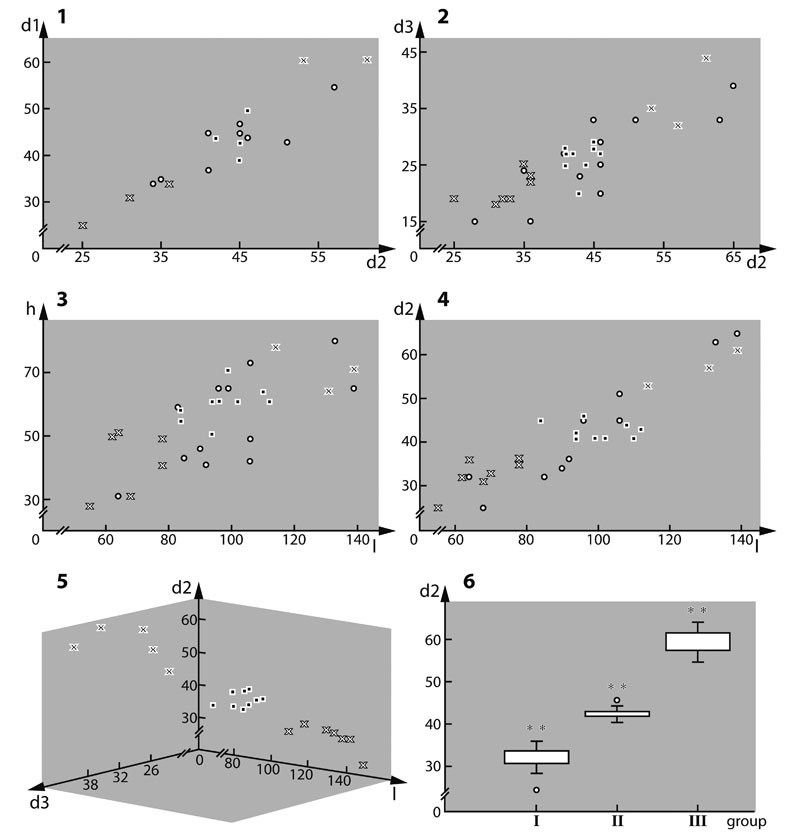
FIGURE 3. Red-blue stereo images of very well preserved specimens processed after the method of Haug J.T. et al. (2009b). 3.1. Best preserved specimens of size class III (top; collection number AUGD 12449B [AUGD omitted in the following]), II (middle; 12448A), and I (bottom; 12449E) respectively; all three specimens to the same scale. 3.2. 12449C. 3.3. 12449A02. 3.4. 12449A10. 3.5. 12449A04. Abbreviations: atl = antennula; cs = cephalic shield; la = labrum; sl = setule; sp = spine; ss = surrounding seta; st = seta. Other abbreviations as before.
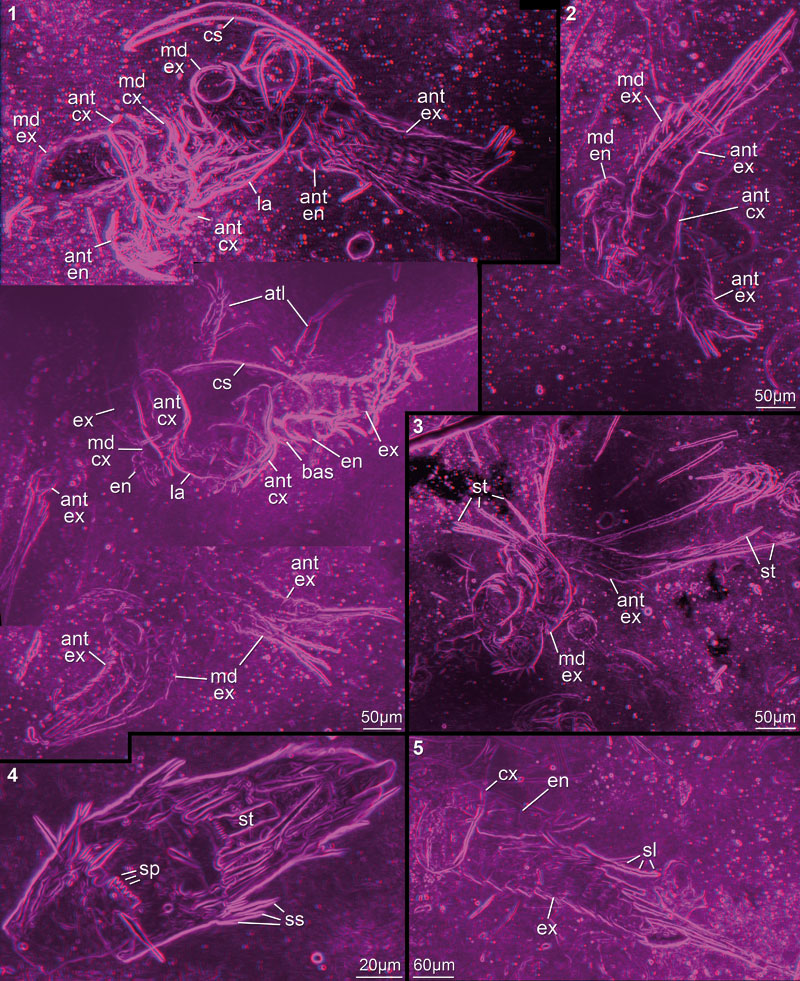
FIGURE 4. Red-blue stereo images of well preserved specimens processed after the method of Haug J.T. et al. (2009b). This method is disadvantageous here, as strongly contrasted areas of the matrix, so-called "curtains" (ct), appear to cut the animal into two halves. 4.1. 12449A25. 4.2. 12449A05. 4.3. 12449A08.
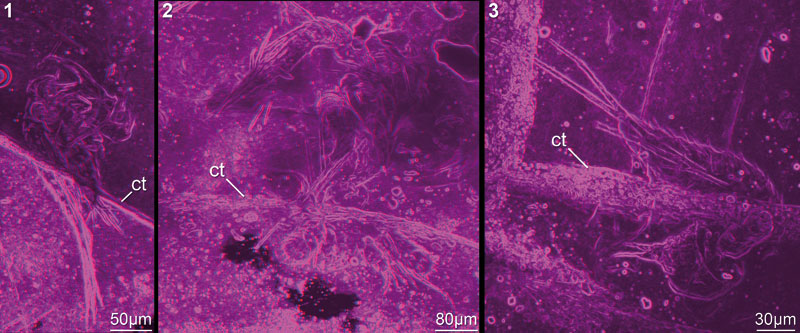
FIGURE 5. Red-blue stereo images of very well preserved specimens. 5.1–2. Rather complete specimens indicating position of appendages in relation to each other. 5.1. 12449B. 5.2. 12448A. 5.3–5. Mandible with many details preserved; 12449A01. 5.3. Entire specimen. 5.4. Terminal setae with setules and visible subdivision (arrow). 5.5. Spines on basipod and coxa. Abbreviations as before.
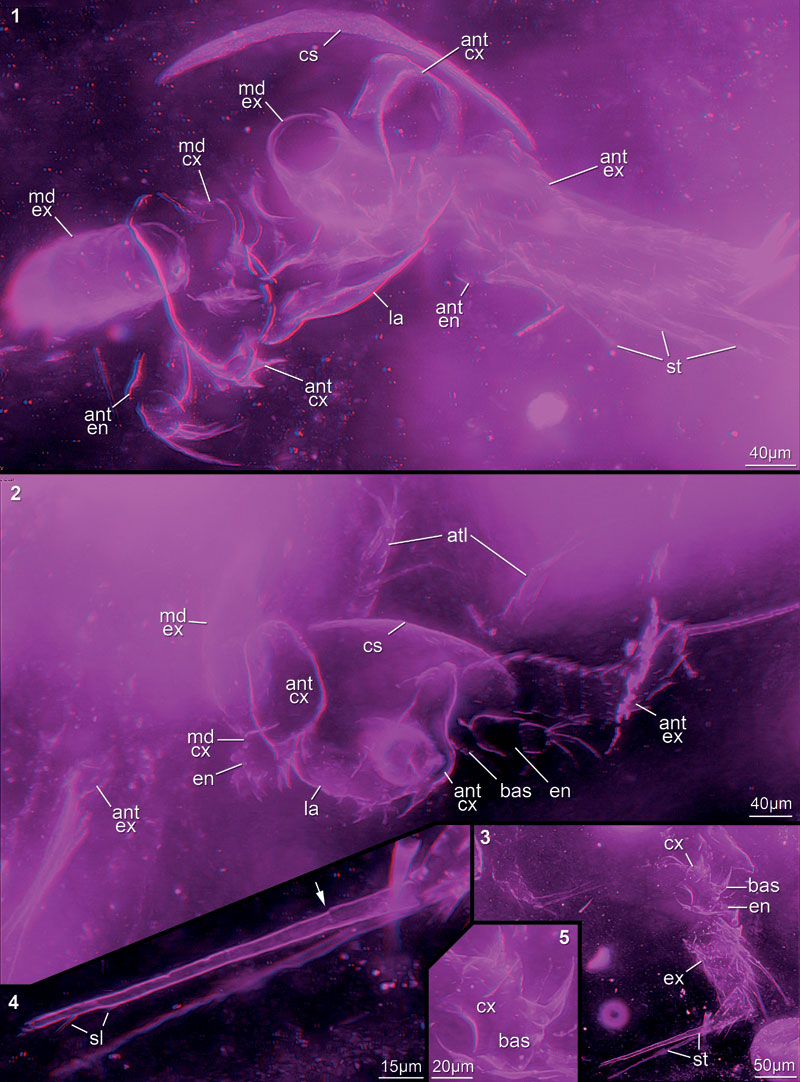
FIGURE 6. Red-blue stereo images of well preserved specimens mainly representing several appendages of one nauplius. 6.1. 12448B. 6.2. 12449C. 6.3. 12449A25. 6.4. 12449A02. 6.5. 12449A03. 6.6. 12449E. 6.7–8. 12449A04. 6.8. Minute setules preserved. Abbreviations as before.

FIGURE 7. Red-blue stereo images of well preserved specimens, but in cloudy-appearing matrix. 7.1. 12445D. 7.2. 12449A20. 7.3. 12449A18. 7.4. 12451F. 7.5. 12447C. 7.6. 12447A. 7.7. 12449A22. Abbreviations as before.
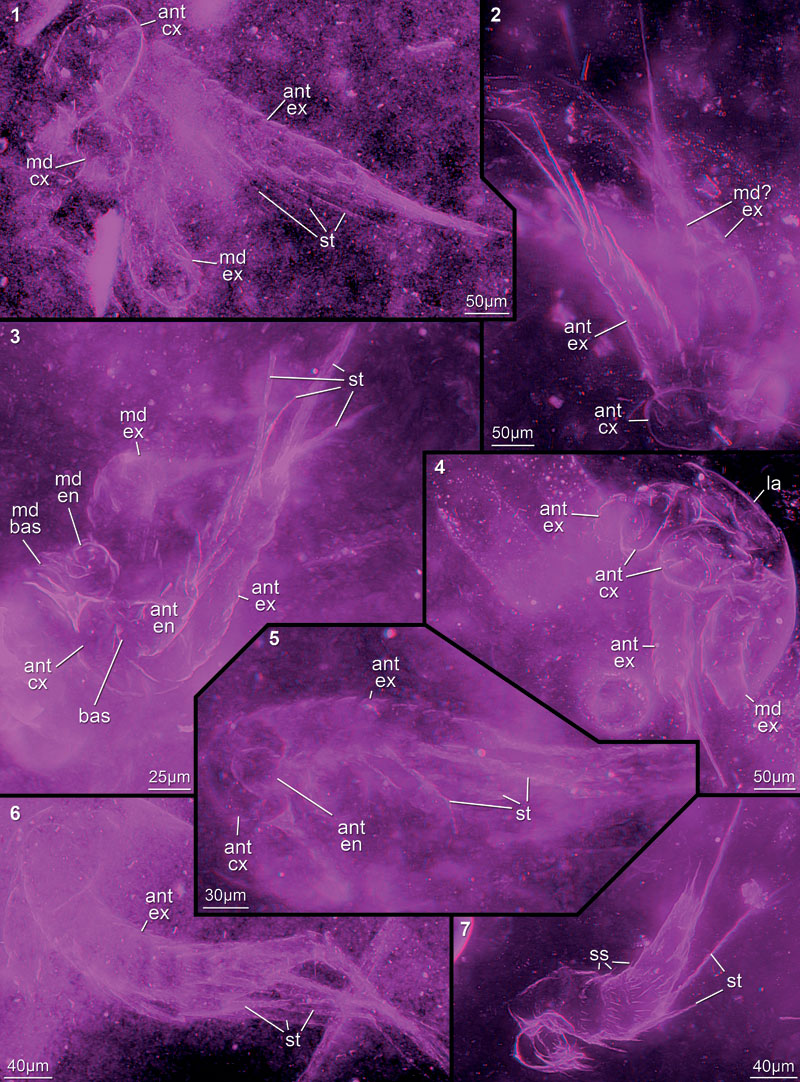
FIGURE 8. Red-blue stereo images of well preserved specimens, but matrix contains many particles or "curtains" (supposedly cracks). 8.1. 12449A05. 8.2. 12449A06. 8.3. 12449A08. 8.4. 12449A16. 8.5. 12449A09. 8.6. 12449A14. 8.7. 12449A19. 8.8. 12447B. 8.9. 12454C. Abbreviations as before.

FIGURE 9. Red-blue stereo images of appendage fragments, mainly parts of exopods (9.1–8), but also coxae (9.9–11). 9.1. 12452B. 9.2. 12449A21. 9.3. 12449A10. 9.4. 12451B. 9.5. 12447F. 9.6. 12449G. 9.7. 12454F. 9.8. 12442E. 9.9. 12454D; supposed coxa in unusual preservation. 9.10. 12451C. 9.11. 12449A15. Abbreviations as before.
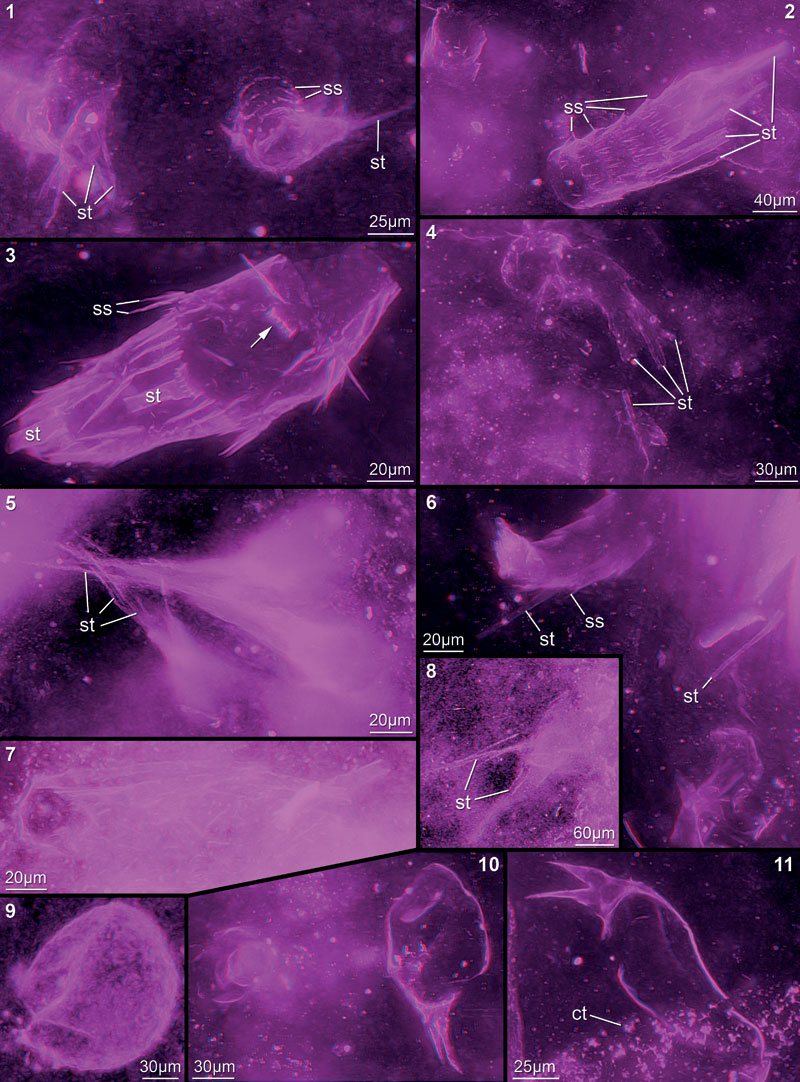
FIGURE 10. Red-blue stereo images of appendage fragments in cloudy-appearing matrix areas, probably all representing exopod fragments. 10.1. 12442B. 10.2. 12445A. 10.3. 12447E. 10.4. 12447D. 10.5. 12444B. 10.6. 12448E. 10.7. 12448F. 10.8. 12449A23. 10.9. 12454G. 10.10. 12454I. 10.11. 12454B. 10.12. 12454H. 10.13. 12451D. Abbreviations as before.

FIGURE 11. Red-blue stereo images of rather faintly preserved specimens, but still several details visible. 11.1. 12454E. 11.2. 12442D. 11.3. 12448G. 11.4. 12454A. 11.5. 12451A. 11.6. 12445F. 11.7. 12449A17. 11.8. 12449A11. 11.9. 12449A07. Abbreviations as before.

FIGURE 12. Red-blue stereo images of faintly preserved specimens; still assignment to here presented nauplius larvae possible due to preserved details. 12.1. 12451G. 12.2. 12449D. 12.3. 12449A13. 12.4. 12448C. 12.5. 12449F. 12.6. 12449A12. 12.7. 12452A. 12.8. 12449A24. 12.9. 12451E. Abbreviations as before.
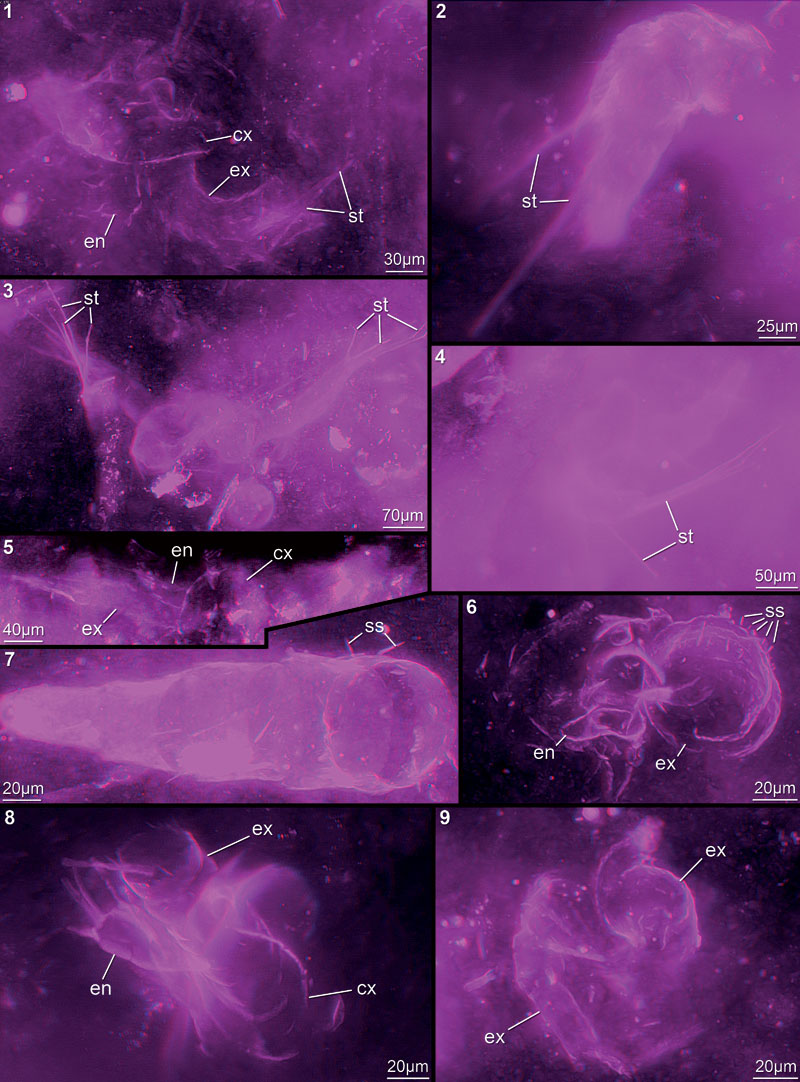
FIGURE 13. Red-blue stereo images of faintly preserved specimens in cloudy-appearing or particle-contaminated matrix areas. 13.1. 12442A. 13.2. 12444A. 13.3. 12442C. 13.4. 12448D. 13.5. 12445E. 13.6. 12448H. 13.7. 12445B. 13.8. 12447G. 13.9. 12442F. Abbreviations as before.
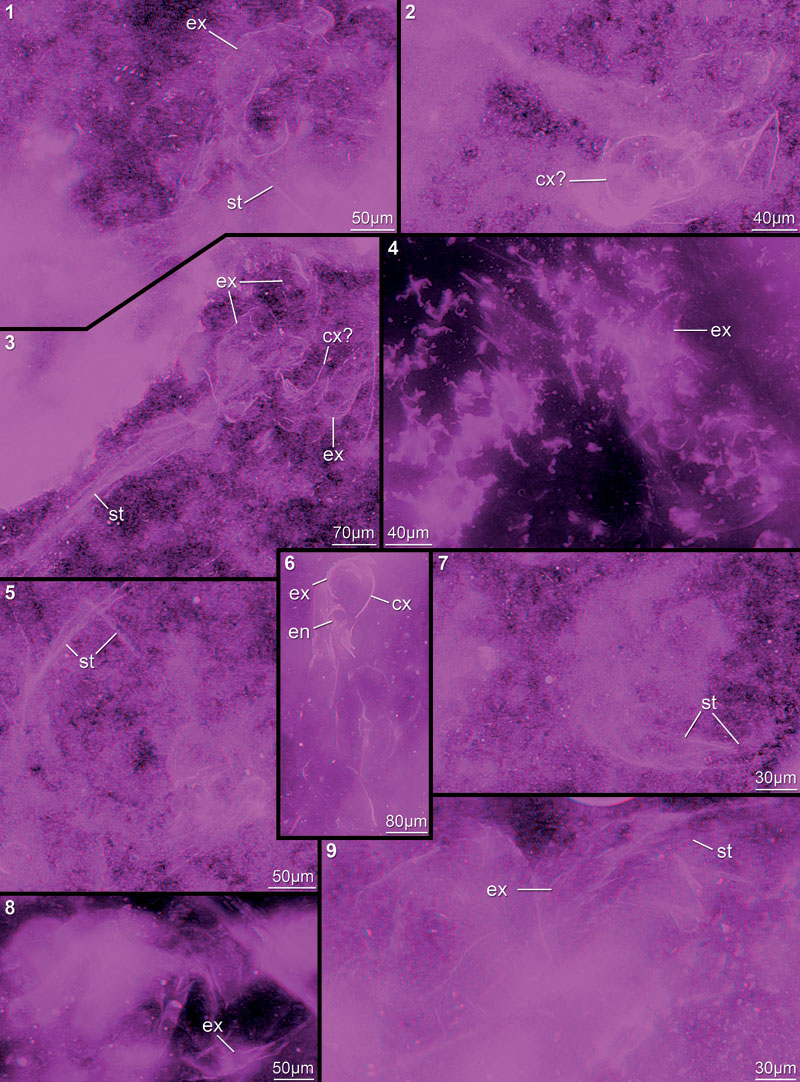
FIGURE 14. Red-cyan stereo image of 3D reconstruction of the here described nauplius in ventral view. Body in dark grey to indicate lacking data.

FIGURE 15. Fossil material for comparisons. 15.1. Marria walcotti Ruedemann, 1931 (USNM 83485A), a graptolite colony formerly erroneously interpreted as a crustacean nauplius; photographed under polarised light. 15.2. Dissected Holocene representative of the copepod species Enhydrosoma gariensis Gurney, 1930 (NHM In 51842); documented under transmitted light microscope with differential interference contrast (DIC).

Exceptionally preserved nauplius larvae from the Devonian Windyfield chert, Rhynie, Aberdeenshire, Scotland
Plain Language Abstract
Seventy-nine specimens of a eucrustacean larva (= nauplius) from the Windyfield chert lagerstätte from NE Scotland, which lies near the famous Rhynie chert site, have been investigated. The larvae have already been briefly described before and interpreted as fairy shrimps (Anostraca), a group within Eubranchiopoda, which includes, e.g., tadpole shrimps (Notostraca), clam shrimps ("Conchostraca") and water fleas (Cladocera). Since the larvae are preserved in three dimensions, we could record stacks of images of each specimen under a transmitted light microscope, from which we subsequently produced stereo images. Measurements of different body parts and statistical analyses led to the detection of three size classes, each with exactly the same morphology. Based on our new data, we reject the interpretation that these larvae are fairy shrimp larvae. They are not even eubranchiopod larvae, because the latter have a third limb (= mandible) with only one ramus, while the larvae described here have a biramous mandible. It cannot be excluded that they belong to a species, which derived from the lineage leading towards the Eubranchiopoda, i.e., before Eubranchiopoda had evolved. However, affinities to Maxillopoda are likewise possible, a group that includes, e.g., barnacles, copepods and seed shrimps. There are even morphological similarities to representatives of the thecostracan lineage (barnacles and relatives), but since all known living representatives of this group are fully marine, the interpretation is difficult to substantiate.
Resumen en Español
Larvas nauplio excepcionalmente preservadas en los sílex de Windyfield (Devónico, Rhynie, Aberdeenshire, Escocia)
Hemos examinado una gran cantidad de larvas nauplio excepcionalmente bien preservadas procedentes del yacimiento de sílex de Windyfield (Devónico inferior, Rhynie, NE de Escocia), situado a 600 m del conocido lagerstätte de sílex de Rhynie. Dos de los ejemplares habían sido ya mencionados en una descripción anterior del material de Windyfield y asignados a crustáceos anostráceos. De los 79 ejemplares disponibles se obtuvieron series de imágenes (de hasta 302 imágenes simples por cada ejemplar) mediante microscopía de luz transmitida. Posteriormente, las series de imágenes fueron proyectadas para obtener imágenes estereoscópicas de cada ejemplar. El análisis morfológico llevado a cabo para detectar posibles estadios de crecimiento reveló la existencia de tres conjuntos diferenciados. Esos conjuntos podrían corresponder a clases de tamaños, pero no hemos podido establecer ninguna diferencia en cuanto al número de elementos de los apéndices, sedas o estructuras similares entre los miembros de las distintas clases de tamaños. Se confirma la naturaleza eucrustácea de las larvas, pero los nuevos datos no apuntan hacia afinidades anostráceas. Teniendo en cuenta la especial morfología de los apéndices, en particular la presencia de mandíbulas birrámeas, las larvas no parecen ser de eubranquiópodos, cuya mandíbula carece de rama exterior o exópodo. Posiblemente, las larvas pertenecen a una especie derivada desde más abajo del nivel evolutivo de los Eubranchiopoda, aunque una afinidad con los maxilópodos también es posible y estaría apoyada igualmente por los datos morfológicos. Más precisamente, la especie podría ser un representante del linaje de los tecostráceos, aunque el ambiente de depósito, correspondiente a un medio efímero de agua dulce, y el conteto paleogeográfico continental no apoyan esta interpretación dado que todos los representantes actuales de los Thecostraca son marinos.
Palabras clave: Eucrustacea; fósil; ontogenia; Branchiopoda; Maxillopoda; modelo tridimensional
Traducción: Miguel Company
Résumé en Français
Préservation exceptionnelle de larves de nauplius dans la silice dévonienne de Windyfield, Rhynie, Aberdeenshire, Écosse
Nous avons examiné un grand nombre de larves de nauplius provenant de la silice dévonienne de Windyfield, Rhynie, NE Écosse, située a 600m de la célèbre localité lagerstätte de Rhynie chert. Deux spécimens ont déjà été mentionnés brièvement dans une précédente description du matériel de Windyfield, supposés appartenir à des crustacés anostraca. A partir des 79 spécimens disponibles, des piles d'images ont été enregistrées à l'aide d'un microscope optique à lumière transmise, allant jusqu'à 302 images individuelles pour chaque spécimen. Par la suite, les piles d'images on été projetées afin d'obtenir des images stéréo de chaque spécimen. L'analyse morphométrique faite dans le but de détecter d'éventuels stades de croissances a révélé trois différents ensembles. Ces derniers peuvent représenter des classes de tailles, mais nous n'avons pas pu reconnaître de différences dans le nombre d'appendices, setae, ou structures similaires entre les représentants de ces différentes classes de tailles. La nature eucrustacée des larves est confirmée, mais les nouvelles données ne donnent pas d'indices quant à l'affinité anostracée. A la lumière de la morphologie spéciale des appendices, particulièrement la possession de mandibules biramées, les larves ne semblent même pas être des eubranchiopodes, qui ont une mandibule sans ramus externe, l'exopode. Les larves pourraient appartenir à une espèce, qui a dérivée depuis un stade évolutif antérieur aux Eubranchiopoda, mais les affinités avec les maxillopodes sont aussi possibles et également supportées par les données morphologiques. Plus précisément l'espèce pourrait être une représentante d'une lignée de Thecostraca, mais les environnements de dépôts éphémères d'eaux douces et la paléogéographie continentale ne supportent pas cette interprétation puisque tous les représentants actuels de Thecostraca sont marins.
Mots clés: Eucrustacea; fossile; ontogénie; Branchiopoda; Maxillopoda; modèle 3D
Translator: Olivier Maridet
Deutsche Zusammenfassung
Außergewöhnlich gut erhaltene Nauplius-Larven aus dem devonischen Windyfield-Chert, Rhynie, Aberdeenshire, Schottland
Zahlreiche außergewöhnlich gut erhaltene Nauplius-Larven aus dem oberdevonischen Windyfield-Chert, Rhynie, Nordost-Schottland, wurden untersucht. Die Fundstelle befindet sich in 600 m Entfernung von der berühmten Rhynie-Chert-Lagerstätte. Zwei der Exemplare waren bereits in einer früheren Beschreibung des Windyfield-Materials kurz erwähnt und damals als anostrake Krebse gedeutet worden. Von den 79 vorhandenen Exemplaren wurden jeweils Bildstapel mit Durchlichtmikroskopie mit bis zu 302 Einzelbildern pro Stapel aufgenommen. Diese Stapel wurden projiziert um Stereobilder von jedem Exemplar zu erhalten. Mit einer morphometrischen Analyse wurden drei verschiedene Gruppen identifiziert. Diese Gruppen repräsentieren vermutlich Größenklassen; jedoch konnten keinerlei Unterschiede in der Zahl der Beinelemente, Seten oder ähnlicher Strukturen zwischen den Vertretern der verschiedenen Größenklassen gefunden werden. Die Zuordnung der Larven zu den Eucrustacea konnte bestätigt werden; die Daten deuten jedoch nicht auf eine Zugehörigkeit zu Anostraca hin. Aufgrund ihrer Beinmorphologie, vor allem wegen des Besitzes biramer Mandibeln, scheinen die Larven keine Eubranchiopoden zu sein, da letzteren der Außenast (= Exopodit) der Mandibel fehlt. Möglicherweise gehören die Larven zu einer Art, welche sich unterhalb des evolutionären Niveaus der Eubranchiopoda abgespalten hat. Auch eine Zugehörigkeit zu den Maxillopoda wäre möglich und wird durch die morphologischen Daten gleichermaßen unterstützt. Genauer gesagt könnte die Art ein Vertreter der Linie der Thecostraca sein. Allerdings wird diese Interpretation durch die Ablagerungsbedingungen in ephemeren Süßgewässern und die kontinentale Paläogeographie nicht unterstützt, da alle heute lebenden Thecostracen marin sind.
Schlüsselwörter: Eucrustacea; Fossil; Ontogenese; Branchiopoda; Maxillopoda; 3D-Modell
Translators: Authors
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024