

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
TABLE 1. Coefficients of Variation (CV) of OPC values for Hoploscaphites spedeni and H. nebrascensis whole shells and separated body chambers and phragmocones of both sexual dimorphs. Note the higher CV for H. spedeni macroconch body chambers and the much smaller CV for H. nebrascensis macroconch body chambers. This reduction in variation may indicate strong selectivity on the body chamber from H. spedeni to subsequent H. nebrascensis.
|
Species |
Dimorph |
n |
CV Whole Shell |
CV Body Chamber |
CV Phragmocone |
|
H. spedeni |
|||||
|
macroconch |
19 |
26.58 |
34.92 |
26.00 |
|
|
microconch |
20 |
24.94 |
28.61 |
28.35 |
|
|
H. nebrascensis |
|||||
|
macroconch |
5 |
14.05 |
11.02 |
31.7 |
|
|
microconch |
5 |
26.16 |
22.95 |
38.80 |
TABLE 2. Comparisons and results of the Mann-Whitney U test for equality of median surface-to-planimetric area ratios. Comparisons were made of body chambers and phragmocones between species and dimorphs of Hoploscaphites. Note that whole shell comparisons were not conducted due to the prohibitively large number of cells per raster. Analyses calculated statistically significant differences at the 95% confidence limit between the median area ratios for all specimens compared. Also note that for the H. spedeni macroconch body chambers, the most rotund, ornate specimens did not necessarily have the largest median cell values. This observation suggests that large changes may occur in late ontogenetic stages within this species.
|
Comparisons |
Specimen 1 Median Surface-to-Planimetric Area Ratio |
Specimen 2 Median Surface-to-Planimetric Area Ratio |
U* |
p (95%) |
|
H. spedeni macro body chamber: |
||||
|
typical vs rotund (ornate) |
1.105 |
1.101 |
200.95 |
< 0.01 |
|
typical vs compressed (smooth) |
1.105 |
1.056 |
25.33 |
< 0.01 |
|
H. spedeni macro phragmocone: |
||||
|
typical vs rotund (ornate) |
1.091 |
1.141 |
30.81 |
< 0.01 |
|
typical vs compressed (smooth) |
1.091 |
1.092 |
4.91 |
< 0.01 |
|
H. spedeni and H. nebrascensis macro body chamber: |
||||
|
rotund H. spedeni vs lowest H. nebrascensis |
1.101 |
1.054 |
44.61 |
< 0.01 |
|
rotund H. spedeni vs highest H. nebrascensis |
1.101 |
1.089 |
13.15 |
< 0.01 |
|
H. spedeni and H. nebrascensis macro phragmocone: |
||||
|
rotund H. spedeni vs lowest H. nebrascensis |
1.141 |
1.070 |
48.43 |
< 0.01 |
|
rotund H. spedeni vs highest H. nebrascensis |
1.141 |
1.109 |
23.17 |
< 0.01 |
|
H. nebrascensis macro body chamber: |
||||
|
lowest vs highest |
1.054 |
1.089 |
204.08 |
< 0.01 |
|
H. nebrascensis macro phragmocone: |
||||
|
lowest vs highest |
1.070 |
1.109 |
9.70 |
< 0.01 |
|
H. spedeni micro body chamber: |
||||
|
typical vs rotund (ornate) |
1.086 |
1.139 |
32.68 |
< 0.01 |
|
typical vs compressed (smooth) |
1.086 |
1.059 |
28.65 |
< 0.01 |
|
H. spedeni micro phragmocone: |
||||
|
typical vs rotund (ornate) |
1.120 |
1.129 |
9.56 |
< 0.01 |
|
typical vs compressed (smooth) |
1.120 |
1.092 |
21.27 |
< 0.01 |
|
H. spedeni and H. nebrascensis micro body chamber: |
||||
|
rotund H. spedeni vs lowest H. nebrascensis |
1.139 |
1.046 |
66.41 |
< 0.01 |
|
rotund H. spedeni vs highest H. nebrascensis |
1.139 |
1.100 |
26.30 |
< 0.01 |
|
H. spedeni and H. nebrascensis micro phragmocone: |
||||
|
rotund H. spedeni vs lowest H. nebrascensis |
1.129 |
1.073 |
42.12 |
< 0.01 |
|
rotund H. spedeni vs highest H. nebrascensis |
1.129 |
1.127 |
7.15 |
< 0.01 |
|
H. nebrascensis micro body chamber: |
||||
|
lowest vs highest |
1.046 |
1.100 |
41.50 |
< 0.01 |
|
H. nebrascensis micro phragmocone: |
||||
|
lowest vs highest |
1.073 |
1.127 |
33.16 |
< 0.01 |
APPENDIX. Three-dimensional rendering of an ammonoid specimen (YPM 23122). This video created in 123D Catch (Autodesk, Inc., 2013) demonstrates the accuracy of this photogrammetric software in creating 3D digital representations. Despite errors or holes around the lower shell margin or keel, the exposed, topside lateral surface of the shell is accurately recreated; features such as ornamentation and flank convexity are realistic in their digital representation in comparison to the actual specimen from which the model was made (refer to Figure 3 for sample photographs used to generate this model). Click on image to run animation.

FIGURE 1. Hoploscaphites spedeni from the Late Cretaceous (Maastrichtian) Western Interior exhibiting intraspecific variation in both dimorphs. Scale bars indicate 1 cm. 1.1, Compressed macroconch (YPM 27162) with subdued ribs and exhibiting only the umbilicolateral and ventrolateral tubercles. 1.2, Typical macroconch (YPM 27160) with better defined ribs, stronger umbilicolateral and ventrolateral tubercles, and numerous randomly distributed tubercles on the recurved hook. 1.3, Rotund macroconch (YPM 23122) with broad ribs, three to four rows of flank tubercles on phragmocone, and randomly distributed tubercles on the recurved hook. 1.4, Compressed microconch (YPM 23199) with broad ribs on the phragmocone, strong umbilicolateral tubercles, and broad ventrolateral tubercles. 1.5, Typical microconch (YPM 23732) with subdued umbilicolateral tubercles, stouter body chamber, and reduced ventrolateral tubercles on recurved hook. 1.6, Ornate microconch (YPM 23699) exhibiting mid-flank row of body chamber tubercles, closely spaced umbilicolaterals, and closely spaced phragmocone ribs. Asterisk (*) indicates the base of the adult body chamber. Modified from Landman and Waage (1993).

FIGURE 2. Hoploscaphites nebrascensis, presumed descendant and sister taxon to H. spedeni, from the Late Cretaceous (Maastrichtian) Western Interior. Scale bars indicate 1 cm. 2.1, 2.2 and 2.3, Macroconchs (YPM 23146, YPM 23145 and YPM 23147, respectively) exhibit a relatively less variable morphology with five to seven rows of flank tubercles from phragmocone through body chamber, a compressed shell, and high whorl height. 2.4, 2.5 and 2.6, Microconchs (YPM 23198, YPM 23687 and YPM 23697, respectively) are consistently compressed like their respective macroconchs, but lack ventrolateral tubercles on the recurved hook, are more variable in the degree of tuberculation ranging from multiple rows of flank tubercles to none, and sometimes contain two rows of closely approximated umbilicolateral tubercles. Asterisk (*) indicates the base of the adult body chamber. Modified from Landman and Waage (1993).

FIGURE 3. Sample photographs used in the creation of 3D digital models (here, a rotund Hoploscaphites spedeni macroconch, YPM 23122). After positioning the specimen and verifying that the dorso-ventral plane was parallel to the table, 140 to 200 photographs were collected at various heights every 10° around the specimen (using angles marked off along the rim of the pan as a guide), to ensure full coverage around each ammonoid.

FIGURE 4. Various orientations of a triangular irregular network (TIN) 3D model for specimen YPM 23122 generated within ArcGIS 10 (ESRI, Inc., 2011) after post-modeling processing within the free, open-source software package, MeshLab (Cignoni et al., 2011). After models were created in 123D Catch (Autodesk, Inc., 2013), they were imported into MeshLab to remove extraneous background scatter (see Appendix 1) and to cut the specimen along the dorso-ventral plane. The XYZ points were exported from MeshLab and then imported into ArcGIS 10, where a TIN surface, like that shown here, was generated for each ammonoid specimen. Note that the accuracy of the digital model was not significantly reduced; however, slight differences are present between the original model made by 123D Catch and the TIN surface generated by ArcGIS 10 from the XYZ points, particularly in the shape of the surficial features (i.e., tubercles) of the phragmocone.

FIGURE 5. Examples of triangular irregular network (TIN) models exhibiting aspect (slope direction) used to generate Orientation Patch Count (OPC) metric, or the number of discrete patches of different aspect classes (e.g., North, Northeast, East, etc.). The color wheel in the center illustrates the orientation of aspect observed on the ammonoids. A feature class containing the number of discrete aspect patches was extracted for each ammonoid, with the total number of patches representing the OPC value. Because the default class breaks for TIN surfaces within ArcGIS 10 contain two North classes (337.5° to 359.9° and 0° to 22.5°), the number of North patches is inherently inflated. Likewise, some specimens have small, seemingly randomly assigned patches around the shell margins, possibly from errors produced in generating the TIN surface. However, all specimens were equally subject to the same problems, suggesting no bias was present to influence the results. 5.1, Hoploscaphites spedeni macroconch (YPM 23122). 5.2, H. spedeni microconch (YPM 23199). 5.3, H. nebrascensis macroconch (YPM 23144). 5.4, H. nebrascensis microconch (YPM 23195).
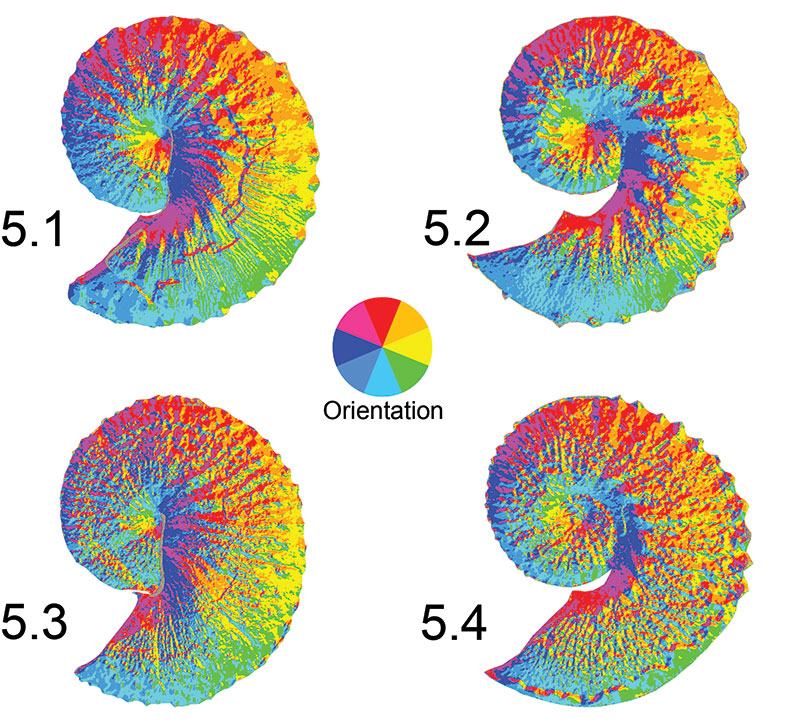
FIGURE 6. Examples of raster models showing the surface-to-planimetric (3D to 2D) area ratio. The legend in the center indicates high values (very steep slope) to low values (near horizontal slope). The mean and median cell values of each raster were extracted and used in comparisons. Rasterization was necessary to conduct map algebra in ArcGIS 10 (ESRI, Inc., 2011). While rasterization does result in reduced resolution of fine features (e.g., ornamentation), the resolution of flank convexity was maintained. 6.1, Hoploscaphites spedeni macroconch (YPM 23122). 6.2, H. spedeni microconch (YPM 23199). 6.3, H. nebrascensis macroconch (YPM 23144). 6.4, H. nebrascensis microconch (YPM 23195).
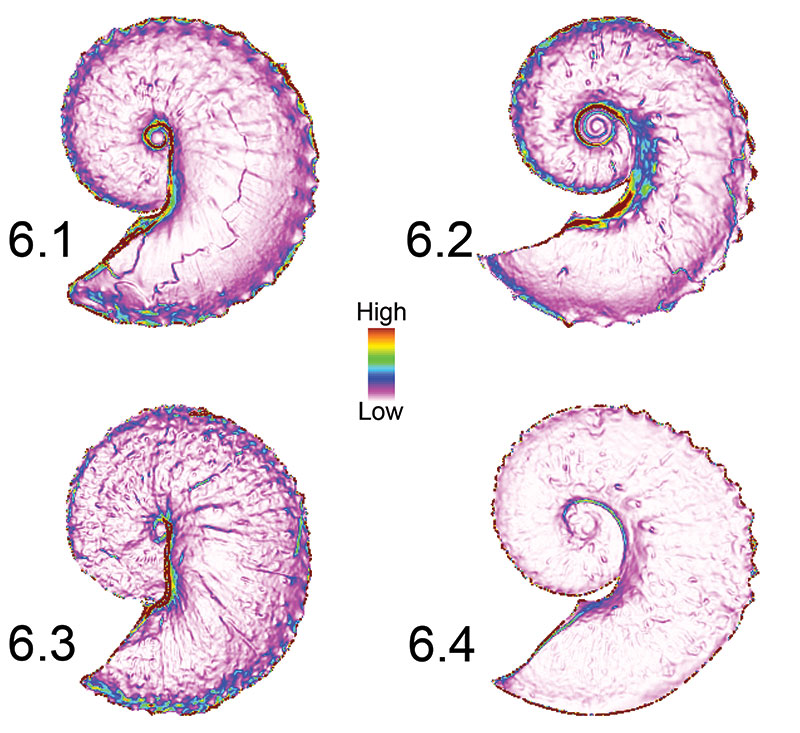
FIGURE 7. Surface aspect maps. The color wheel in the center illustrates the orientation of aspect observed on the ammonoids. Plotting surface aspect (slope direction) allows visualization of ornamentation across the lateral surface of Hoploscaphites. 7.1, H. nebrascensis macroconch (YPM 23147). A section (black box) of the lateral surface of the shell near the base of the body chamber is magnified to demonstrate that tubercles (topographically, a round hill) can show nearly all aspect classes. Note the circled tubercle within the box; it exhibits all aspect classes. Therefore, ammonoids with many tubercles should have higher OPC values. The H. nebrascensis specimen shown here has an OPC value of 57,759 patches. 7.2, H. spedeni macroconch (YPM 44402). A section (black box) of the lateral surface of the shell at the point of recurvature on the body chamber is magnified to demonstrate that ribs (topographically, an extended ridge) show alternating aspect classes. Therefore, ammonoids with few tubercles (i.e., rib dominated) should have lower OPC values. The H. spedeni specimen shown here has an OPC value of 35,678 patches.
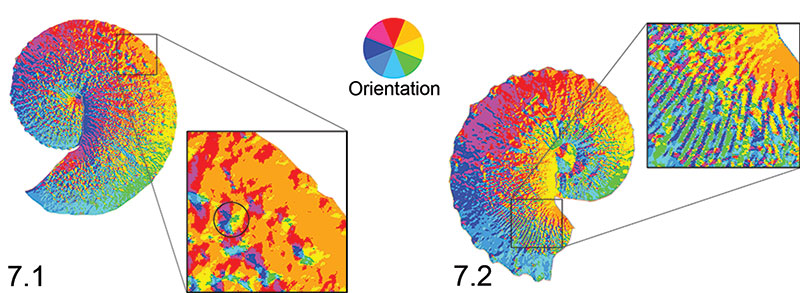
FIGURE 8. Reduced major axis regression of Orientation Patch Count (OPC) vs. maximum shell length (LMAX, mm). OPC values are positively correlated with maximum shell length, indicating that the largest specimens, which tend to be the most tuberculate, have the largest OPC values.
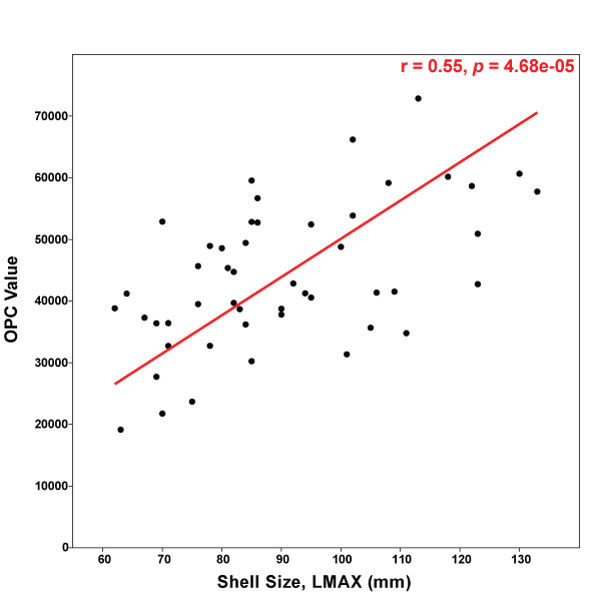
FIGURE 9. Frequency histograms of OPC value for whole shells, body chambers and phragmocones, comparing two dimorphs. Macroconchs (gray bars, blue normality curve) and microconchs (black bars, red normality curve) were compared for both Hoploscaphites spedeni and H. nebrascensis separately. Student's t and F tests were applied to compare the distributions of these data. 9.1, Frequency histogram of OPC value for H. spedeni whole shells. Macroconchs had a significantly larger mean OPC than microconchs at 95% confidence. 9.2, Frequency histogram of OPC value for H. spedeni body chambers; no statistically significant differences between dimorphs. 9.3, Frequency histogram of OPC value for H. spedeni phragmocones; no significant differences between dimorphs. 9.4, Frequency histogram of OPC value for H. nebrascensis whole shells; no significant differences between dimorphs. 9.5, Frequency histogram of OPC value for H. nebrascensis body chambers; no significant difference between dimorphs. 9.6, Frequency histogram of OPC value for H. nebrascensis phragmocones; no significant difference between dimorphs.
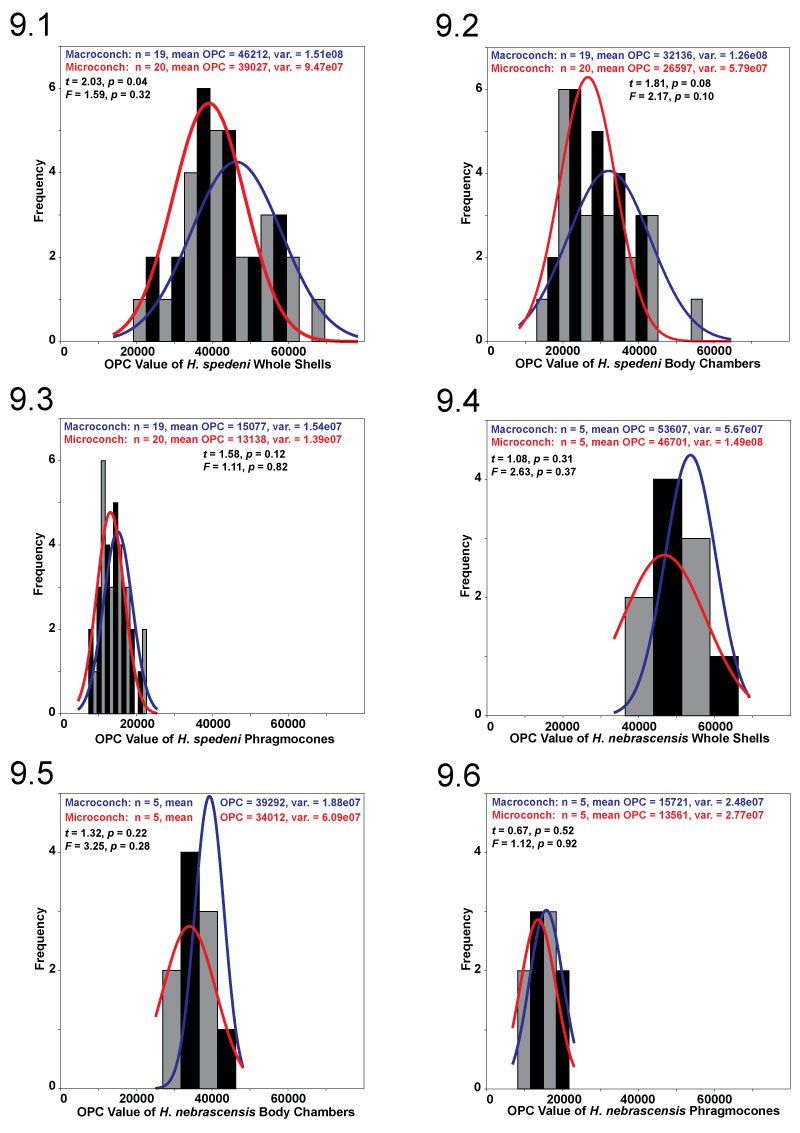
FIGURE 10. Frequency histograms of OPC value for whole shells, body chambers, and phragmocones, comparing two Hoploscaphites species. Macroconchs for both Hoploscaphites spedeni (gray bars, blue normality curve) and H. nebrascensis (black bars, red normality curve) were compared to each other separately from microconchs of H. spedeni (gray bars, blue normality curve) and H. nebrascensis (black bars, red normality curve). Student's t and F tests were applied to compare the distributions of these data. No statistically significant differences were determined at 95% confidence in all comparisons. 10.1, Frequency histogram of OPC value for H. spedeni and H. nebrascensis macroconch whole shells. 10.2, Frequency histogram of OPC value for H. spedeni and H. nebrascensis macroconch body chambers. 10.3, Frequency histogram of OPC value for H. spedeni and H. nebrascensis macroconch phragmocones. 10.4, Frequency histogram of OPC value for H. spedeni and H. nebrascensis microconch whole shells. 10.5, Frequency histogram of OPC value for H. spedeni and H. nebrascensis microconch body chambers. 10.6, Frequency histogram of OPC value for H. spedeni and H. nebrascensis microconch phragmocones.
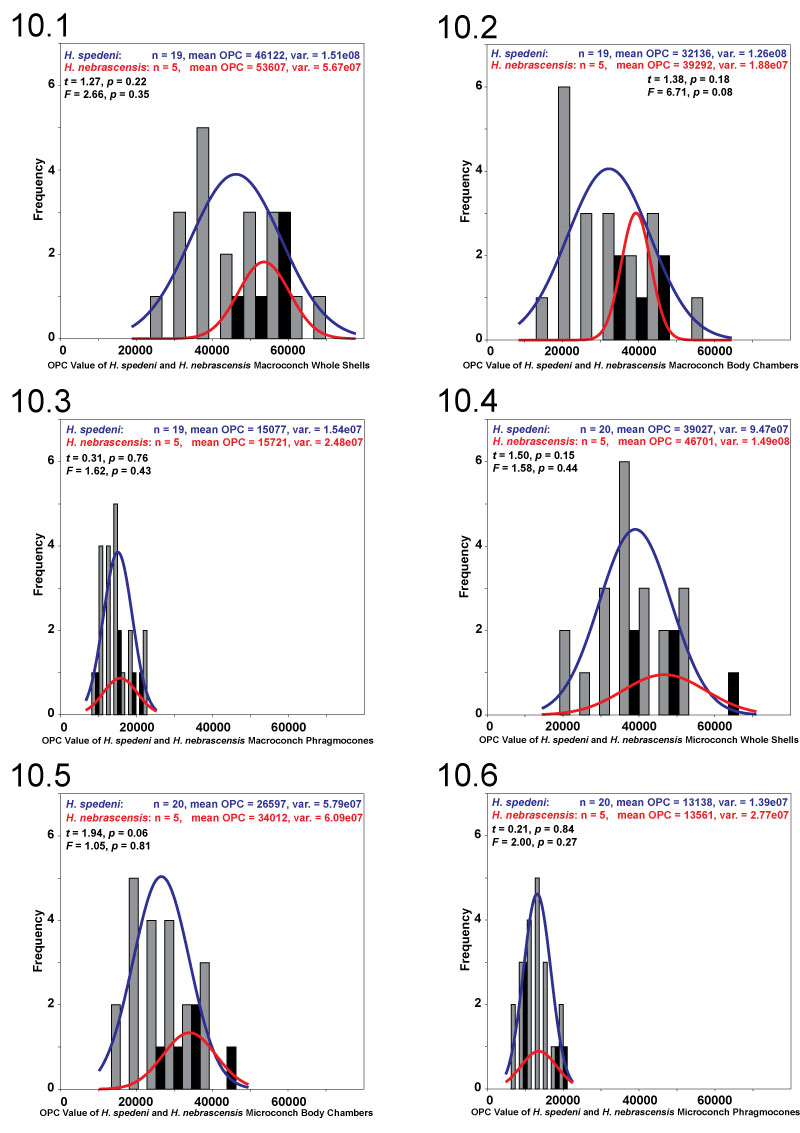
FIGURE 11. Surface-to-planimetric (3D to 2D) area ratio. Large-scale topographic features are most likely being quantified by this metric. These features may include the degree of shell compression or flank convexity, the size and overall shape of the ventrolateral tubercles around the margins of the shell, some combination of both, or some other shell feature(s). 11.1, Hoploscaphites spedeni macroconch (YPM 23124) with the typical morphotype exhibiting flank convexity or depression (more "High" 3D to 2D area ratio coloration around the margins). 11.2, H. nebrascensis macroconch (YPM 23145) exhibiting the relatively compressed, or flattened, flank shape (less "High" 3D to 2D area ratio coloration around the margins and uniformity in overall coloration).
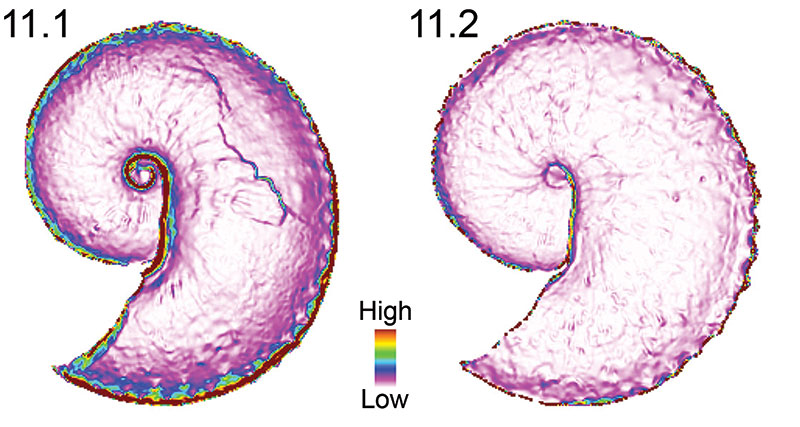
FIGURE 12. Reduced major axis regression of the square root of the surface-to-planimetric (3D to 2D) area ratio mean and median cell values versus different morphometric variables. 12.1, No statistically significant linear correlation between the square root of the mean area ratios and OPC values. 12.2, No statistically significant linear correlation between the square root of the median area ratios and OPC values. The lack of correlation between these two independent surface ruggedness indices indicates that these two metrics are measuring two different shell characters or aspects of shell characters. 12.3, No statistically significant linear correlation between the square root of the mean area ratios and aperture width (mm). 12.4, No statistically significant linear correlation between the square root of the median area ratios and aperture width (mm). 12.5, No statistically significant linear correlation between the square root of the mean area ratios and width of the shell at the ultimate septum (mm). 12.6, No statistically significant correlation between the square root of the median area ratios and width of the shell at the ultimate septum (mm).
 Mathew J. Knauss
Mathew J. Knauss
Department of Earth Sciences
University of California
Riverside
1242 Geology Building
Riverside, California, 92521
USA
mknau001@ucr.edu
Mathew Knauss received both his B.S. in Geology (Paleobiology Specialization) and M.S. in Geology at Bowling Green State University (Bowling Green, Ohio, USA). He is currently a Ph.D. student in the Hughes Laboratory at the University of California, Riverside. His research interests focus on developmental plasticity, intraspecific variability, and functional morphology of both ammonoids and trilobites. His current research involves quantifying morphological and ontogenetic variability in scaphitid ammonoids of the Late Cretaceous Western Interior Sea Way, and understanding the evolution of trilobite enrollment, specifically focusing on the microstructure and biomechanics of the exoskeleton and the differences in coaptative devices that aid in enrollment.

 Margaret M. Yacobucci
Margaret M. Yacobucci
Department of Geology
Bowling Green State University
190 Overman Hall
Bowling Green, Ohio, 43403-0218
USA
mmyacob@bgsu.edu
Peg (Margaret M.) Yacobucci received her A.B. in Geophysical Sciences from the University of Chicago, and her Ph.D. in Earth and Planetary Sciences from Harvard University. She is currently Associate Professor of Geology at Bowling Green State University. Her research centers on understanding the evolution of fossil cephalopods, especially Cretaceous ammonoids, through analyses of phylogenetic history, morphological and developmental plasticity, and paleobiogeography. She currently serves as the Paleontological Society’s Education and Outreach Coordinator (2010-2016), and as an Associate Editor for the Journal of Paleontology and Palaios.
Geographic Information Systems technology as a morphometric tool for quantifying morphological variation in an ammonoid clade
Plain Language Abstract
Ammonoids are known for their anatomical variation within and between closely related species. However, few researchers have successfully quantified this variation due to the complexity of ammonoid characters. Ammonoid shells are three-dimensional (3D) structures with a variety of anatomical features. Characters such as shell shape and ornamentation often covary and change throughout growth. It is difficult to describe, quantify and compare these features using standard paleontological techniques. Geographic information systems (GIS), computer software widely used by geoscientists to visualize, process and collect new geographic or spatial information, may allow for quantification of these complex shell characters. Here, we present a new GIS-based method for visualizing and quantifying variation within and between ammonoid species in their full 3D form.
After making 3D digital models of specimens belonging to the ancestral and more variable ammonoid species Hoploscaphites spedeni and its less variable descendant H. nebrascensis from the Late Cretaceous Western Interior of North America, two terrain roughness indices were applied to the surface to quantify ornamentation and shell shape differences. The first metric, called Orientation Patch Count (OPC), quantified the number of discrete patches on the surface of the shell that have different slope directions. This metric allowed for quantification of the amount of ornamentation. For example, ammonoids with many bumps on their shells had many different slope direction patches, and hence, had a higher OPC value than those specimens with fewer bumps. The second metric, the ratio of the actual curved surface (3D) area and the equivalent flat (2D) area of the shell, quantified changes in slope across the surface of the shell. The results of this metric were more difficult to interpret but most likely reflect how curved the edge of the shell was.
The results of this GIS-based analysis support the interpretation that H. nebrascensis had juvenile-like ornamentation (i.e., bumps) in its adult form compared to its ancestor, H. spedeni, which reduced or lost most of its bumps throughout its late growth stages before reaching full maturity. Additionally, the H. nebrascensis macroconchs (the presumed females) became less variable in their conch morphology compared to the H. spedeni macroconchs. However, microconchs (the presumed males) of H. nebrascensis retained the variation of their ancestor, H. spedeni.
These spatial analyses demonstrate the power that GIS holds for quantifying complex fossil forms and addressing questions related to evolutionary relationships and shell growth. Further refinement of this method plus the addition of other GIS-based spatial analyses may allow for researchers to visualize and quantify different, complex fossil anatomies in their full 3D form.
Resumen en Español
Tecnología de Sistemas de Información Geográfica (SIG) como una herramienta morfométrica para cuantificar la variación morfológica en un clado de amonoideos
Los amonoideos son conocidos por su variación morfológica intraespecífica e interespecífica durante la ontogenia, en particular en la forma de la concha y en la ornamentación. Muchas de las características covarían y los elementos individuales son difíciles de homologar, lo que hace inapropiadas, para la cuantificación de estas morfologías complejas, tanto las descripciones cualitativas como las herramientas morfométricas más ampliamente utilizadas. Sin embargo, los análisis espaciales llevados a cabo en un entorno SIG (Sistema de Información Geográfica) permiten la cuantificación y visualización de la forma global de la concha. A continuación, presentamos una metodología basada en el SIG, en la cual la variación de las características de la concha es considerada para evaluar los patrones evolutivos en un clado de amonoideos del Cretácico tardío del Mar Interior Occidental de América del Norte. Investigamos el ancestral y más variable amonites heteromorfo de la familia Scaphitidae Hoploscaphites spedeni y su descendiente menos variable H. nebrascensis. Se crearon modelos digitales en tres dimensiones (3D) de las superficies laterales de las conchas fósiles usando un software fotogramétrico y se importaron las reconstrucciones en un entorno SIG. Se utilizó el número de áreas de aspecto discreto y las proporciones de la superficie de área planimétrica (3D a 2D) de la superficie lateral como índices de la rugosidad del terreno. Estos análisis espaciales en 3D muestran los rangos morfológicos superpuestos de H. spedeni y H. nebrascensis en la ornamentación a pesar de las diferencias en la compresión de la concha. Además, la diana para el cambio evolutivo en este clado se encuentra en la cámara del cuerpo de la macroconcha, la cual se vuelve menos variable a lo largo del desarrollo de la macroconcha desde H. spedeni hasta su transformación a H. nebrascensis; sin embargo, las microconchas mantienen la variación ancestral a lo largo de la ontogenia. Estos análisis geoespaciales no solo cuantificaron satisfactoriamente la variación en morfologías complejas, sino que también demostraron la versatilidad de este método para afrontar cuestiones relacionadas con la ontogenia y la filogenia.
Palabras clave: amonoideos; variación intraespecífica; ontogenia; SIG; morfometría
Traducción: Enrique Peñalver
Résumé en Français
La technologie des systèmes d'information géographique comme outil morphométrique pour quantifier la variation morphologique dans un clade ammonoïde
Les Ammonites sont connus pour leur variation morphologique intraspécifique et interspécifique par l'ontogenèse, en particulier la forme de la coquille et l'ornementation. De nombreuses caractéristiques covarient et certains éléments individuels sont difficiles à homologuer, ce qui fait des descriptions qualitatives et des outils morphométriques largement utilisés inappropriés pour quantifier ces morphologies complexes. Cependant, des analyses spatiales, menées dans un environnement de système d'information géographique (SIG), permettent de la quantification et la visualisation des formes de coquille mondiale. Ici, nous présentons une méthodologie basée sur le SIG dans laquelle la variation des caracteristiques de la coquille est déterminée pour évaluer les tendances évolutive dans un clade ammonoïde de la voie maritime intérieure de l'ouest de l'Amérique du Nord du Crétacé supérieur. Nous étudions l'ammonoïde scaphitide hétéromorphe ancestrale et plus variable, Hoploscaphites spedeni, et son moins variable descendant H. nebrascensis. Nous avons créé des modèles numériques en trois dimensions (3D) de surfaces latérales de coquilles fossiles, en utilisant un logiciel de photogrammétrie et importé les reconstructions dans un environnement SIG. Nous avons utilisé le nombre de parcelle à aspect discrètes et les rapports de surface- planimétrique (3D à 2D) de la zone de la surface latérale comme indices de rugosité du terrain. Ces analyses spatiales 3D montrent le chevauchement des gammes morphologiques de H. spedeni et H. nebrascensis dans l'ornementation, malgré les différences de compression de la coquille. En outre, l'objectif de changement évolutif dans ce clade réside dans la chambre du corps macroconque, qui devient moins variable à travers le développement de H. spedeni vers H. nebrascensis dans leurs macroconques; toutefois, les microconques conservent la variation des ancêtres à travers l'ontogenèse. Ces analyses géospatiales quantifies non seulement la variation des morphologies complexes avec succès, mais a également démontré la polyvalence de cette méthode pour aborder des questions liées à l'ontogenèse et la phylogenèse.
Mots-clés: ammonites; variation intraspécifique; ontogenèse; SIG; morphométriques
Translator: Kenny J. Travouillon
Deutsche Zusammenfassung
Geographische Informationssytem-Technologie als morphometrisches Werkzeug zur Quantifizierung morphologischer Variation bei einer Ammoniten-Klade
Ammoniten sind für ihre intraspezifische und interspezifische morphologische Variation während der Ontogenese bekannt, besonders was Gehäusegestalt und Ornamentierung angeht. Viele Merkmale co-variieren und individuelle Elemente sind schwer zu homologisieren, weshalb qualitative Beschreibungen und weit verbreitete morphometrische Werkzeuge für eine Quantifizierung dieser komplexe Morphologie ungeeignet sind. Jedoch erlauben räumliche Analysen, die in einer geographischen Informationssytem (GIS) Umgebung durchgeführt wurden, die Quantifizierung und Visualisierung einer globalen Gehäusegestalt. Wir präsentieren hier eine GIS-basierte Methode bei der die Variation der Gehäusemerkmale abgeschätzt wurde, um die evolutionären Muster in einer Ammonitenklade aus dem spätkretazischen Western Interior Seaway von Nordamerika zu evaluieren. Wir untersuchen den älteren und stärker variablen scaphitiden heteromorphen Ammoniten Hoploscaphites spedeni und seinen weniger variablen Nachkommen H. nebrascensis. Mittels photogrammetrischer Software generierten wir dreidimensionale (3D) digitale Modelle der lateralen Gehäuseoberflächen des Fossils und importierten die Rekonstruktionen in eine GIS Umgebung. Wir verwendeten die Anzahl der getrennten Seitenflecken und das Verhältnis von Oberfläche zu planimetrischer Ebene (3D zu 2D) der lateralen Oberflächen als Rauheitsindizes. Diese 3D räumlichen Analysen demonstrieren die Überlappung der morphologischen Reichweiten von H. spedeni und H. nebrascensis im Bereich der Ornamentierung trotz der Unterschiede bei der Gehäusekompression. Zusätzlich liegt das Ziel für eine evolutionäre Veränderung in dieser Klade in der Gehäusekammer des Macroconchen, das bei den Macroconchen während der Entwicklung von H. spedeni zum nachfolgenden H. nebrascensis weniger variabel wird. Die Microconche jedoch behalten ihre angestammte Variation während der Ontogenese bei. Diese geospatialen Analysen quantifizierten nicht nur erfolgreich Variation bei komplexer Morphologie, sondern zeigten ebenso die vielseitige Verwendbarkeit dieser Methode auf, wie Fragen zu Ontogenese und Phylogenie angegangen werden können.
Schlüsselwörter: Ammoniten; intraspezifische Variation; Ontogenese; GIS; Morphometrie
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024