

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
Appendix 1. List of included UCMP and LACM specimens from Thousand Creek, Nevada. Abbreviations: #, number; R, right; L, left; I., Ilingoceros; S., Sphenophalos; LM, medial length; TD, distal thickness; TI, intermediate thickness; TP, proximal thickness; LL, lateral length; WD, distal width; WI, intermediate width; LI, intermediate length. (PDF format)
Appendix 2. Results of the discriminant function analysis on the set of measurements of Antilocapra americana. Abbreviations: M, male; F, female; R, right; L, left. All specimens are from the museum of Vertebrate Zoology (MVZ). (PDF format)
FIGURE 1. Photographs of fossil antilocaprid horns. 1.1, Ilingoceros schizoceras; 1.2, I. alexandrae; 1.3, Sphenophalos nevadanus.

FIGURE 2. Illustration of idealized antilocaprid astragalus, indicating the eight dimensions used in this analysis, following DeGusta and Vrba (2003). 2.1, anterior view; 2.2, lateral View; 2.3, medial view. Abbreviations: LM= medial length; LI= intermediate length; LL= lateral length; TD= distal thickness; TI= intermediate thickness; TP= proximal thickness; WD= distal width; WI= intermediate width.
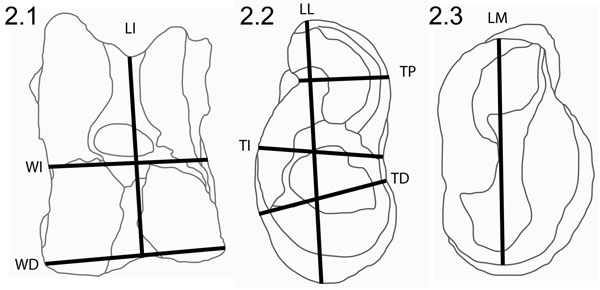
FIGURE 3. Principal components plots of Thousand Creek Antelope data. 3.1, PC1 versus PC2; 3.2, PC2 versus PC3.
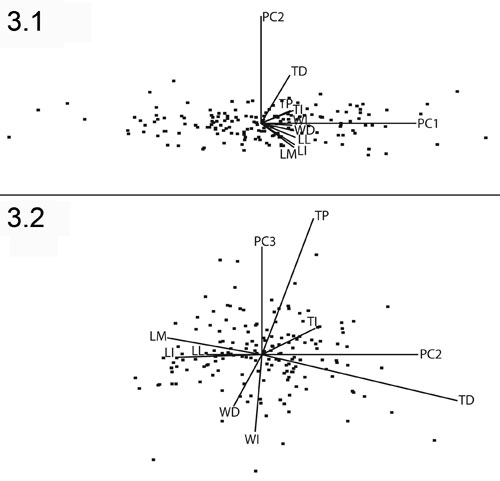
FIGURE 4. Comparison of values of V* for the Thousand Creek antilocaprids, Texoceros guymonensis, and Antilocapra americana. Vertical bars represent 95% confidence intervals.
TABLE 1. Summary statistics of astragali samples.
|
n |
mean |
std dev |
S-W1 |
V2 |
V *3 |
SV* 4 |
||
|
Thousand Creek |
LM |
190 |
27.83 |
1.71 |
n.s. |
0.0614 |
0.0615 |
0.0032 |
|
TD |
211 |
13.99 |
1.25 |
n.s. |
0.0893 |
0.0895 |
0.0044 |
|
|
TI |
216 |
15.30 |
1.27 |
n.s. |
0.0830 |
0.0831 |
0.0040 |
|
|
TP |
212 |
11.06 |
0.97 |
n.s. |
0.0877 |
0.0878 |
0.0043 |
|
|
LL |
198 |
30.51 |
2.05 |
n.s. |
0.0672 |
0.0673 |
0.0034 |
|
|
WD |
193 |
18.62 |
1.59 |
0.0153 |
0.0854 |
0.0855 |
0.0044 |
|
|
WI |
201 |
17.71 |
1.60 |
n.s. |
0.0903 |
0.0905 |
0.0046 |
|
|
LI |
215 |
23.88 |
1.58 |
n.s. |
0.0662 |
0.0662 |
0.0032 |
|
|
Antilocapra americana |
LM |
26 |
34.15 |
1.24 |
n.s. |
0.0363 |
0.0367 |
0.0052 |
|
TD |
24 |
16.70 |
1.02 |
n.s. |
0.0611 |
0.0617 |
0.0092 |
|
|
TI |
25 |
18.81 |
0.84 |
n.s. |
0.0447 |
0.0451 |
0.0066 |
|
|
TP |
26 |
13.90 |
0.63 |
n.s. |
0.0453 |
0.0458 |
0.0065 |
|
|
LL |
25 |
36.72 |
1.44 |
0.0039 |
0.0392 |
0.0396 |
0.0058 |
|
|
WD |
26 |
21.91 |
0.80 |
0.0428 |
0.0365 |
0.0369 |
0.0053 |
|
|
WI |
26 |
22.39 |
1.04 |
n.s. |
0.0464 |
0.0469 |
0.0067 |
|
|
LI |
25 |
28.93 |
1.06 |
0.0357 |
0.0366 |
0.0370 |
0.0054 |
|
|
Texoceros guymonensis |
LM |
67 |
22.75 |
1.02 |
n.s. |
0.0448 |
0.0450 |
0.0039 |
|
TD |
67 |
11.51 |
0.96 |
n.s. |
0.0834 |
0.0837 |
0.0074 |
|
|
TI |
67 |
12.49 |
0.91 |
n.s. |
0.0729 |
0.0731 |
0.0064 |
|
|
TP |
67 |
9.39 |
0.65 |
n.s. |
0.0692 |
0.0695 |
0.0061 |
|
|
LL |
67 |
24.95 |
1.19 |
n.s. |
0.0477 |
0.0479 |
0.0042 |
|
|
WD |
67 |
15.23 |
0.90 |
n.s. |
0.0591 |
0.0593 |
0.0052 |
|
|
WI |
65 |
14.46 |
1.10 |
n.s. |
0.0761 |
0.0764 |
0.0068 |
|
|
LI |
65 |
19.92 |
1.08 |
n.s. |
0.0542 |
0.0544 |
0.0048 |
1 Significance of deviation from normal distribution using a Shapiro-Wilk test.
2 Coefficient of Variation.
3 V corrected for sample size.
4 Standard Error of V*.
TABLE 2. Intra-operator error for astragalar dimensions.
|
Dimension |
Difference |
Difference |
|
LM |
0.12 |
0.4 |
|
TD |
0.65 |
4.5 |
|
TI |
0.36 |
2.3 |
|
TP |
0.14 |
1.3 |
|
LL |
0.11 |
0.4 |
|
WD |
0.15 |
0.8 |
|
WI |
0.24 |
1.3 |
|
LI |
0.12 |
0.5 |
TABLE 3. First three principal components of Thousand Creek antilocaprid astragali.
|
PC1 |
PC2 |
PC3 |
|
|
Size variation |
Size-independent variation in shape |
||
|
Eigenvalue |
6.5276 |
0.4219 |
0.3504 |
|
Percent |
81.5945 |
5.2741 |
4.3795 |
|
Eigenvectors |
|||
|
LM |
0.3651 |
-0.3738 |
0.0965 |
|
TD |
0.3178 |
0.7765 |
-0.2646 |
|
TI |
0.3590 |
0.2098 |
0.1506 |
|
TP |
0.3284 |
0.2069 |
0.7848 |
|
LL |
0.3764 |
-0.2173 |
0.0021 |
|
WD |
0.3631 |
-0.1106 |
-0.2944 |
|
WI |
0.3450 |
-0.0260 |
-0.4419 |
|
LI |
0.3692 |
-0.3322 |
-0.0120 |
TABLE 4. Summary statistics of Bovid astragali.
|
Species |
mean |
sd |
V |
V* |
s.e. V* |
V* - 1kCr V* |
p |
|
|
Aepyceros melapus |
LM |
35.04 |
1.410 |
0.0402 |
0.0412 |
0.0103 |
-1.61 |
0.109 |
|
n=10 |
TD |
16.53 |
1.036 |
0.0627 |
0.0642 |
0.0164 |
-1.49 |
0.138 |
|
TI |
19.20 |
0.894 |
0.0466 |
0.0477 |
0.0120 |
-2.79 |
0.006 |
|
|
TP |
13.76 |
0.660 |
0.0480 |
0.0492 |
0.0124 |
-2.94 |
0.004 |
|
|
LL |
37.42 |
1.580 |
0.0422 |
0.0433 |
0.0109 |
-2.11 |
0.036 |
|
|
WD |
21.78 |
0.900 |
0.0413 |
0.0424 |
0.0106 |
-3.75 |
0.000 |
|
|
WI |
21.86 |
0.903 |
0.0413 |
0.0424 |
0.0106 |
-4.16 |
0.000 |
|
|
LI |
28.94 |
1.195 |
0.0413 |
0.0423 |
0.0106 |
-2.15 |
0.032 |
|
|
Antidorcas marsupialis |
LM |
28.16 |
1.637 |
0.0581 |
0.0599 |
0.0174 |
-0.09 |
0.930 |
|
n=8 |
TD |
13.58 |
1.193 |
0.0879 |
0.0906 |
0.0269 |
0.04 |
0.967 |
|
TI |
15.73 |
1.043 |
0.0663 |
0.0684 |
0.0199 |
-0.72 |
0.470 |
|
|
TP |
11.16 |
0.938 |
0.0841 |
0.0867 |
0.0256 |
-0.04 |
0.966 |
|
|
LL |
30.25 |
1.739 |
0.0575 |
0.0593 |
0.0172 |
-0.46 |
0.647 |
|
|
WD |
18.79 |
1.132 |
0.0602 |
0.0621 |
0.0180 |
-1.26 |
0.209 |
|
|
WI |
18.81 |
1.499 |
0.0797 |
0.0822 |
0.0242 |
-0.34 |
0.736 |
|
|
LI |
23.66 |
1.349 |
0.0570 |
0.0588 |
0.0170 |
-0.43 |
0.669 |
|
|
Gazella granti |
LM |
33.80 |
1.340 |
0.0396 |
0.0406 |
0.0102 |
-1.96 |
0.052 |
|
n=10 |
TD |
17.04 |
0.837 |
0.0491 |
0.0503 |
0.0127 |
-2.91 |
0.004 |
|
TI |
19.08 |
0.842 |
0.0441 |
0.0452 |
0.0114 |
-3.15 |
0.002 |
|
|
TP |
13.33 |
0.943 |
0.0707 |
0.0725 |
0.0186 |
-0.80 |
0.424 |
|
|
LL |
36.74 |
1.503 |
0.0409 |
0.0419 |
0.0105 |
-2.30 |
0.023 |
|
|
WD |
21.95 |
0.920 |
0.0419 |
0.0430 |
0.0108 |
-3.65 |
0.000 |
|
|
WI |
20.79 |
0.794 |
0.0382 |
0.0391 |
0.0098 |
-4.75 |
0.000 |
|
|
LI |
28.65 |
1.154 |
0.0403 |
0.0413 |
0.0103 |
-2.30 |
0.022 |
|
|
Tragelaphus scriptus |
LM |
33.6 |
2.729 |
0.08122 |
0.08291 |
0.01937 |
1.09 |
0.277 |
|
n=12 |
TD |
15.23 |
1.495 |
0.09816 |
0.10021 |
0.02373 |
0.44 |
0.658 |
|
TI |
18.03 |
1.597 |
0.08857 |
0.09042 |
0.02125 |
0.34 |
0.735 |
|
|
TP |
13.41 |
1.25 |
0.09321 |
0.09516 |
0.02244 |
0.32 |
0.748 |
|
|
LL |
35.73 |
3.106 |
0.08693 |
0.08874 |
0.02083 |
1.02 |
0.311 |
|
|
WD |
21.23 |
1.903 |
0.08964 |
0.0915 |
0.02152 |
0.27 |
0.785 |
|
|
WI |
19.59 |
1.987 |
0.10143 |
0.10354 |
0.02458 |
0.52 |
0.603 |
|
|
LI |
28.96 |
2.539 |
0.08767 |
0.0895 |
0.02102 |
1.10 |
0.274 |
TABLE 5. Comparison of V*, following Sokal and Braumann (1980).
|
V*1-V*2 1 |
S V*1-V*2 2 |
df 3 |
t 4 |
p |
||
|
Thousand Creek |
LM |
0.0249 |
0.0061 |
214 |
4.0560 |
<0.0001 |
|
TD |
0.0277 |
0.0102 |
233 |
2.7131 |
0.0072 |
|
|
TI |
0.0380 |
0.0077 |
239 |
4.9170 |
<0.0001 |
|
|
TP |
0.0420 |
0.0078 |
236 |
5.3637 |
<0.0001 |
|
|
LL |
0.0277 |
0.0067 |
221 |
4.1217 |
<0.0001 |
|
|
WD |
0.0486 |
0.0069 |
217 |
7.0840 |
<0.0001 |
|
|
WI |
0.0436 |
0.0081 |
225 |
5.3675 |
<0.0001 |
|
|
LI |
0.0292 |
0.0063 |
238 |
4.6488 |
<0.0001 |
|
|
Thousand Creek |
LM |
0.0165 |
0.0051 |
255 |
3.2634 |
0.0013 |
|
TD |
0.0057 |
0.0086 |
276 |
0.6687 |
n.s. |
|
|
TI |
0.0100 |
0.0076 |
281 |
1.3144 |
n.s. |
|
|
TP |
0.0183 |
0.0075 |
277 |
2.4535 |
0.0148 |
|
|
LL |
0.0194 |
0.0054 |
263 |
3.5911 |
0.0004 |
|
|
WD |
0.0262 |
0.0068 |
258 |
3.8444 |
0.0002 |
|
|
WI |
0.0141 |
0.0082 |
264 |
1.7181 |
n.s. |
|
|
LI |
0.0118 |
0.0058 |
278 |
2.0317 |
0.0431 |
|
|
T. guymonensis |
LM |
0.0083 |
0.0066 |
91 |
1.2725 |
n.s. |
|
TD |
0.0220 |
0.0118 |
89 |
1.8637 |
n.s. |
|
|
TI |
0.0280 |
0.0092 |
90 |
3.0462 |
0.0030 |
|
|
TP |
0.0237 |
0.0089 |
91 |
2.6512 |
0.0095 |
|
|
LL |
0.0083 |
0.0071 |
90 |
1.1573 |
n.s. |
|
|
WD |
0.0225 |
0.0074 |
91 |
3.0323 |
0.0032 |
|
|
WI |
0.0295 |
0.0096 |
89 |
3.0813 |
0.0027 |
|
|
LI |
0.0174 |
0.0073 |
88 |
2.4009 |
0.0185 |
Differences in coefficients of variation.
2 Standard Error of difference.
3 Degrees of freedom.
4 V*1-V*2 / S V*1-V*2.
Extending the utility of artiodactyl postcrania for species-level identifications using multivariate morphometric analyses
Edward Byrd Davis and Jonathan Jean-Michel Calède
Plain Language Abstract
Can the ankle bones of fossil artiodactyls be used for species-level identifications?
Changes in the diversity and abundance of life through geological time can be best understood when fossil samples have many specimens that have been clearly identified. We are interested in the paleoecology of fossil Antilocaprids, a family of ruminant artiodactyls (related to cows, deer, and giraffes) found only in North America. Today the family is represented only by the pronghorn antelope, Antilocapra americana, the fastest land animal in North America, but in the Miocene (23-5 million years ago) the diversity of fossil antilocaprids approached the diversity of modern antelope in Africa. Species of fossil pronghorn can be identified using the shape of their horncores (the bone support inside the horn), but the rest of their skeletons are often difficult to distinguish. Horncores are rare in fossil assemblages, but bones of the feet are common. If we could identify species from foot bones, we could dramatically increase the total sample size of fossil pronghorn. In fact, if the method we develop here for pronghorn proves successful, it could be applied to other groups of artiodactyls as well, so our results have implications for fossil camels, deer, and many other cloven-hoofed animals. To test whether it is possible to use the size and shape of these foot bones (specifically the ankle bone called the astragalus) to identify species of fossil pronghorn, we analyzed a set of measurements from the ankle bones of present-day African antelope, which have often been considered ecologically similar to the extinct pronghorn species. Our results indicate that species African antelope can be clearly distinguished using their anklebones, so we extended the study to a group of three fossil pronghorn species. The three species, Ilingoceros alexandrae, Ilingoceros schizoceras, and Sphenophalos nevadanus lived about 8 million years ago in a site in northwestern Nevada called Thousand Creek. Like most fossil pronghorn, these species are identified from their distinctive horncores, but previous researchers have suggested they might actually represent bucks and does or adults and fawns from a single species. We measured over 200 pronghorn ankle bones from this Nevada site. Our results indicate that while there is enough variation in anklebone size and shape to reject the hypothesis that they come from only one species, the data do not have a clear enough signal to allow assignment of individual specimens to species. Additionally, our comparison of previous workers’ results on African antelope to ankle bones of modern pronghorn indicates that the pronghorn have a different enough evolutionary history that African antelope may not be a good guide to the ecology of extinct pronghorn antelope.
Glossary:
Antilocaprid: member of the family that includes the modern pronghorn, Antilocapra americana.
Artiodactyl: a member of the order of mammals that have ‘cloven feet’. This includes pigs, deer, cattle, pronghorn, camels, and giraffes, amongst many others.
Astragalus: the bone that connects the foot to the lower leg, the ankle bone.
Bovid: member of the family that includes cattle, goats, and African antelope.
Coefficient of Variation (CV, V): A measurement of the variability in a sample of measurements that is unaffected by the absolute size of the specimens measured. CV should be comparable between samples of mice and elephants.
Discriminant Function Analysis (DFA): A statistical analysis that allows a user to distinguish between two or more kinds of specimens using measurements of those specimens.
Hemphillian: a time period in the late Miocene, from 9 to 5 million years ago.
Horncore: the bone support for a horn, found in bovids (like cattle) and pronghorn antelope.
Postcrania: the parts of a skeleton that are not in the skull.
Podials: the many small bones of the hands and feet.
Resumen en Español
Utilidad de los elementos postcraneales de los artiodáctilos para la identificación a nivel específico mediante el uso de análisis morfométricos multivariantes
Los estudios paleoecológicos son más fiables cuanto mayor sea el tamaño de las muestras analizadas y más fina la escala taxonómica con la que se trabaja. Los elementos postcraneales suelen ser abundantes pero raramente son identificativos a nivel específico, viéndose así reducido el potencial del tamaño de la muestra. En este estudio hemos investigado si los astrágalos de los antilocápridos, abundantes en el Mioceno superior de la Gran Cuenca, pueden servir para la identificación de especies, con lo que se optimizarían los tamaños muestrales. El análisis de los datos bibliográficos existentes sobre bóvidos asiáticos y africanos sugiere que se pueden diferenciar especies a partir de las dimensiones del astrágalo. Para nuestro estudio hemos utilizado tres especies de antilocápridos (Ilingoceros alexandrae, Ilingoceros schizoceras y Sphenophalos nevadanus) del Hemfiliense (~8 Ma) presentes en la Fauna de Thousand Creek del noroeste de Nevada. Estas especies se diferencian unas de otras por el tipo de cornamenta, mientras que las comparaciones previas de su dentición no han mostrado una clara separación entre ellas. Nuestro análisis de más de 200 astrágalos de antilocápridos de Thousand Creek indica que existe variación suficiente como para rechazar en principio la hipótesis de que se trate de una única especie, aunque la distribución no permite la atribución específica de los astrágalos. Combinados con la morfología de las astas, nuestros resultados sugieren que las diferencias en la competición entre los machos y una pequeña divergencia en el tamaño corporal excluirían la posibilidad de que hubiera competencia entre ambos géneros, mientras que la similitud ecológica y/o la existencia de un ancestro común habrían dado lugar a una distribución continua de las dimensiones del astrágalo. Hemos explorado, además, la efectividad de una función discriminante publicada con anterioridad para inferir preferencias ambientales a partir de los astrágalos en los bóvidos africanos. La aplicación de esta función discriminante en Antilocapra no da lugar a resultados satisfactorios, probablemente como consecuencia de las distintas historias evolutivas de antilocápridos y bóvidos.
PALABRAS CLAVE: análisis discriminante; coeficiente de variación; Antilocapridae; Bovidae; hábitat; abundancia.
Traducción: Miguel Company
Résumé en Français
Etendre l’utilité des restes post-crâniens d’artiodactyles pour les identifications au niveau spécifique par l’utilisation d’analyses morphologiques multivariées
Les études paléoécologiques sont d’autant plus performantes avec des données sur l’abondance relative à une échelle taxinomique fine et sur de larges échantillons.
Les éléments post-crâniens sont abondants mais rarement identifiés au niveau spécifique, réduisant potentiellement la taille de l’échantillon. Nous testons si les astragales d’antilocapridés, abondants dans le Miocène supérieur du ‘Great Basin’, peuvent être identifiés au niveau spécifique de façon à augmenter la taille des échantillons. Notre analyse des données de bovidés africains et asiatiques issues de la littérature suggère que les espèces peuvent être distinguées par l’utilisation des dimensions de l’astragale. Pour notre cas d’étude nous utilisons trois espèces d’antilocapridés, Ilingoceros alexandrae, Ilingoceros schizoceras, et Sphenophalos nevadanus de la faune ‘Thousand Creek’ hemphillienne (~8 Ma) du nord-ouest du Nevada. Ces espèces sont diagnostiquées par leurs noyaux de cornes, mais les comparaisons précédentes de leurs dentitions n’ont montré aucune séparation claire entre les espèces. Notre analyse de plus de 200 astragales d’antilocapridés provenant de ‘Thousand Creek’ indique une variabilité suffisamment importante pour exclure l’hypothèse d’une seule espèce, mais la distribution ne permet pas d’assigner chaque astragale à une espèce particulière. Combinée avec la morphologie des noyaux de cornes, nos résultats suggèrent que des différences dans la compétition male-male et une légère différence de taille ont évité la compétition entre les deux genres, alors que la similarité écologique et/ou une ascendance commune ont créé une distribution continue de la dimension des astragales. Les données ne permettent pas confirmer si I. alexandrae et I. schizoceras sont bien des espèces distinctes. Par ailleurs, nous avons testé le domaine d’efficacité d’une fonction discriminante développée à partir des astragales bovidés africains et publiée dans le but d’interpréter les préférences environnementales. L’application de cette fonction discriminante à Antilocapra s’est révélée inefficace, probablement en raison d’histoires évolutives différentes entre antilocapridés et bovidés.
MOTS CLES : analyse discriminante ; CV ; Antilocapridae ; Bovidae ; habitat ; abondance
Translator: Olivier Maridet
Deutsche Zusammenfassung
Ausbau des Nutzwertes von Paarhufer - Postcranialskeletten zur Artniveaubestimmung mit multivariaten morphometrischen Analysen
Paläoökologische Untersuchungen sind am aussagekräftigsten, wenn relative Häufigkeitsangaben in kleinen taxonomische Einheiten und großen Probengrößen vorliegen. Postcranialelemente sind häufig, doch selten können Arten identifiziert werden, was die potentielle Probengröße reduziert. Wir untersuchen ob bei antilocapriden Astragali, die im späten Miozän des Great Basin häufig vorkommen, Arten identifiziert werden können um damit die Probengrößen zu erweitern. Unsere Datenanalysen von afrikanischen und asiatischen Boviden aus der Literatur weisen darauf hin, dass Arten anhand unterschiedlicher Astragalus - Dimensionen zu unterscheiden sein sollten.
Für unsere Fallstudie benutzen wir drei Arten von Antilocapriden, Ilingoceros alexandrae, Ilingoceros schizoceras und Sphenophalos nevadanus aus dem Hemphillian (~8 Ma) der Thousand Creek Fauna im nordwestlichen Nevada. Diese Arten werden anhand des Horns bestimmt, jedoch haben frühere Untersuchungen der Bezahnung keine klare Unterscheidung zwischen den Arten gezeigt. Nach unserer Analyse von mehr als 200 antilocapriden Astragali liegt ausreichend Variation vor, dass die Hypothese von nur einer Art vorläufig verworfen werden kann. Jedoch erlaubt die Verteilung keine Zuordnung von einzelnen Astragali zu einer Art. Kombiniert mit der Hornmorphologie deuten unsere Ergebnisse auf Unterschiede in Männchenkonkurrenz hin. Ein kleiner Größenunterschied trennte die beiden Gattungen, während ökologische Ähnlichkeiten und/oder gemeinsame Vorfahren eine kontinuierliche Verteilung der Astragalus-Dimensionen schufen. Die Daten können nicht auflösen ob I. alexandrae und I. schizoceras unterschiedliche Arten sind. Zusätzlich haben wir den Wirksamkeitsgrad einer publizierten Diskriminanzfunktion sondiert, der entwickelt wurde, um aus Astragali von afrikanischen Boviden deren Umweltpräferenz abzuleiten. Die Anwendung dieser Diskriminanzfunktion auf Antilocapra hat sich als unwirksam erwiesen, was eine Folge der unterschiedlichen evolutionären Entwicklung von Antilocapridae und Bovidae sein dürfte.
SCHLÜSSELWÖRTER: Diskriminanzanalyse; CV; Antilocapridae; Bovidae; Habitat; Häufigkeit
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
Polski Abstrakt
Rozwinięcie zastosowań materiału pozaczaszkowego parzystokopytnych dla identfikacji na poziomie gatunkowym przy użyciu analizy morfometrycznej wielu zmiennych
Badania paleoekologiczne mają największą moc w przypadku, gdy dane o względnej liczebności są dostępne w małej skali taksonomicznej i przy dużych wielkościach prób. Materiał pozaczaszkowy bywa liczny, ale rzadko identyfikowalny do poziomu gatunku, co redukuje potencjalną wielkość próby. Zbadaliśmy, czy kości skokowe widłorogich, liczne w późno-mioceńskich osadach Wielkiej Kotliny, mogą zostać zidentyfikowane do poziomu gatunku, poprawiając tym samym wielkość próby. Nasza analiza danych literaturowych dotyczących afrykańskich i azjatyckich krętorogich pokazuje, że gatunki powinny być oznaczane względem wymiarów na kościach skokowych. W naszym przypadku użyliśmy trzech gatunków widłorogich, Ilingoceros alexandrae, Ilingoceros schizoceras i Sphenophalos nevadanus wieku hemfilliaśkiego (~8 Ma) fauny Thousand Creek (północno-wschodnia Newada). Te gatunki diagnozowane są na podstawie ich rogów, ale wcześniejsze porównania ich uzębienia nie odzwierciedliły granic pomiędzy gatunkami. Nasza analiza ponad 200 kości skokowych widłorogich z Thousand Creek wskazuje, że istnieje wystarczająca zmienność by wstępnie obalić możliwość istnienia tylko jednego gatunku, ale rozkład cech nie umożliwia przypisania konkretnych kości skokowych do gatunków. W porównaniu z morfologią rogów, nasze wyniki sugerują ze różnice we współzawodnictwie między samcami, oraz niewielkie różnice rozmiarów, sprawiały że oba rodzaje nie stanowiły dla siebie konkurencji, podczas gdy podobieństwa w ekologii oraz/lub wspólne pochodzenie skutkowały ciągłym rozkładem wymiarów na kościach skokowych. Dane nie pokazują, czy I. alexandrae i I. schizoceras są odrębnymi gatunkami. Dodatkowo, zbadaliśmy zakres skuteczności opublikowanych funkcji odmienności wypracowanych celem przypisania preferencji środowiskowych do kości skokowych afrykańskich pustorogich. Zastosowanie tych funkcji odmienności do Antilocapra okazało się nieskuteczne, prawdopodobnie w wyniku odmiennych historii ewolucyjnych widłorogich i pustorogich.
Słowa kluczowe: analiza odmienności, CV, Antilocapridae, Bovidae, środowisko, liczebność
Translators: Dawid Mazurek and Robert Bronowicz
Riassunto in Italiano
Tentativo di identificazione specifica attraverso elementi postcraniali negli artiodattili mediante l’utilizzo di analisi morfometriche multivariate
Gli studi paleoecologici sono più significativi quando sono disponibili dati di abbondanza relativa a scala tassonomica più fine e su campioni consistenti. Gli elementi postcraniali sono abbondanti, ma raramente è possibile la loro identificazione a livello specifico; ciò riduce la grandezza potenziale del campione. Abbiamo cercato di verificare se gli astragali degli antilocapridi, abbondanti nel Great Basin durante il Miocene superiore, possono essere identificati a livello specifico, in modo da aumentare la grandezza del campione. Un’analisi dei dati di letteratura riguardanti i bovidi asiatici ed africani suggerisce che le dimensioni dell’astragalo dovrebbero essere discriminanti a livello di specie. Nel nostro caso utilizziamo tre specie di antilocapridi (Ilingoceros alexandrae, Ilingoceros schizoceras e Sphenophalos nevadanus) dell’Hemphilliano (~8 Ma) di Thousand Creek Fauna (Nevada nordo-ccidentale). Tali specie si distinguono sulla base dei nuclei ossei delle corna; analisi comparative della dentizione non hanno evidenziato una chiara separazione fra le specie. Abbiamo analizzato più di 200 astragali di antilocapridi provenienti da Thousand Creek, e ne abbiamo concluso che esiste sufficiente variabilità per scartare l’ipotesi della presenza di un’unica specie, ma non è possibile, sulla base della loro distribuzione, caratterizzare a livello specifico i singoli astragali. I nostri risultati, combinati con la morfologia dei nuclei ossei delle corna, sembrano indicare che le differenze nella competizione tra maschi e una leggera differenza di taglia hanno contribuito a tenere i due generi separati. D’altro canto, similarità ecologiche e/o un’ascendenza comune hanno avuto come risultato una distribuzione continua delle dimensioni degli astragali. Dai dati ottenuti non è possibile discriminare le specie I. alexandrae e I. schizoceras. Abbiamo inoltre verificato l’efficacia di una funzione discriminante (disponibile in letteratura) sviluppata per estrapolare dati ambientali a partire da astragali di bovidi africani. L’applicazione di tale funzione ad Antilocapra si è dimostrata inefficace, probabilmente come conseguenza delle distinte storie evolutive di antilocapridi e bovidi.
Parole chiave: analisi discriminante; CV; Antilocapridae; Bovidae; habitat; abbondanza
Translator: Chiara Angelone
Edward Byrd Davis Department of Geological Sciences and Museum of Natural and Cultural History
Department of Geological Sciences and Museum of Natural and Cultural History
University of Oregon
Eugene, Oregon, 97403–1272
USA
Edward Byrd Davis is the Fossil Collections Manager for the University of Oregon Museum of Natural and Cultural History. He received his B.S. from the University of Tennessee, Knoxville, and his Ph.D. from the University of California, Berkeley. He divides his time between museum curational nightmares, teaching in the UO Geology Department, and paleomammalogy research. His primary research interests lie in trends in macroecology over time and in the evolution of antilocaprid artiodactyls.
Jonathan Jean-Michel Calède
Department of Biology
University of Washington
Seattle, Washington, 98195-1800
USA
Jonathan Jean-Michel Calede is a Ph.D. student in the Department of Biology at the University of Washington. He received his B.S. from the Université Claude Bernard Lyon 1 (France) and his M.S. from the University of Oregon. He is starting his dissertation work focusing on Oligocene community dynamics. His research interests include Cenozoic mammalian paleoecology and community structure.
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024