

 A new name for old bones: A reassessment of Early Jurassic theropod remains from Dorset, England
A new name for old bones: A reassessment of Early Jurassic theropod remains from Dorset, England
Article number: 27.2.a26
https://doi.org/10.26879/1346
Copyright Palaeontological Association, May 2024
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 28 September 2023. Acceptance: 22 April 2024.
ABSTRACT
Among the many terrestrial vertebrate specimens that have been recovered from the Lower Jurassic Blue Lias Formation (Hettangian-Sinemurian) in Dorset, England, there are two partial yet distinctive theropodan hindlimbs that are not currently assigned to any recognised, valid taxon. Originally this material was referred to the early armoured ornithischian taxon Scelidosaurus harrisonii, which had also been recovered from a Lower Jurassic Formation in Southern England. However, this error in assignment was soon realised and corrected in subsequent works, and the material in question reclassified as an indeterminate theropod. More recent works have suggested possible neotheropod or even tetanuran affinities for at least some of this material, but no consensus has yet been reached about its true phylogenetic and taxonomic affinities. The material has never been formally named, even though the material does appear to display a distinct combination of anatomical characteristics. As one of the earliest known Jurassic representatives of Theropoda, understanding this material is important for clarifying the picture of early theropod evolution and biogeographic distribution. This study re-appraises the two specimens, compares, and contrasts them with other known Triassic and Early Jurassic theropods, and tests their possible phylogenetic affinities using three different anatomical datasets: two that are heavily theropod focused and one more general early dinosaur focused. The results of these analyses, in combination with the anatomical comparisons presented herein, suggest that this fragmentary material could represent a distinct, early occurring, and early branching averostran neotheropod. A new genus and species are thus erected for the material.
Matthew G. Baron. BPP University, 142 Uxbridge Rd, Uxbridge, London, W12 8AA, UK. m.baron2@my.bpp.com
Keywords: Dinosauria; Mesozoic; phylogeny; systematics; Theropoda; Blue Lias
Final citation: Baron, Matthew G. 2024. A new genus and species of fossil pseudoscorpion (Arachnida: Pseudoscorpiones) from the Eocene amber of Western India. Palaeontologia Electronica, 27(2):a26.
https://doi.org/10.26879/1346
palaeo-electronica.org/content/2024/5067-on-an-early-jurassic-theropod
Copyright: May 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
ZOOBANK to be supplied by PE Editors
INTRODUCTION
Dinosaur remains are well known from several Lower and Middle Jurassic formations within the United Kingdom, and many of these have a very long history of study - indeed, some of the earliest scientifically described dinosaur specimens were recovered from Jurassic formations in Southern England (Buckland, 1824; Mantell, 1825; Owen, 1842, 1859, 1874).
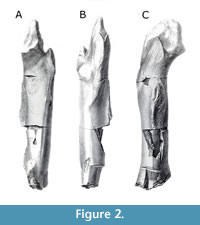
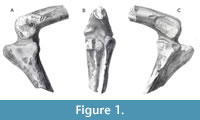 The Blue Lias formation, which is Early Jurassic (Hettangian-Sinemurian) in age (Cope et al., 1980), has yielded a number of dinosaur remains, as well as remains of other terrestrial and marine vertebrates. The ornithodirans are represented in the Blue Lias by the early pterosaur Dimorphodon macronyx (Buckland, 1829; Owen, 1859; Barrett et al., 2008) the theropods Sarcosaurus woodi (Andrews, 1921; Ezcurra et al., 2020) and Dracoraptor hanigani (Martill et al., 2016), as well numerous specimens currently regarded as indeterminate members of Theropoda (see, for example, Ezcurra et al., 2020); amongst these indeterminate theropods are two specimens: the distal portion of a femur, the proximal portions of a tibia and fibula, and a partial femur that is missing its distal and proximal articular surfaces (Figure 1, Figure 2, and Figure 3A-C). These partial hindlimbs, comprising the complete and articulated knee joint (Figure 1A-C, Figure 3A-B), and the partial femur (Figure 2, Figure 3C), were originally assigned to the genus Scelidosaurus by Owen (1859), and later included them within the hypodigm of the newly erected species Scelidosaurus harrisonii (1861).
The Blue Lias formation, which is Early Jurassic (Hettangian-Sinemurian) in age (Cope et al., 1980), has yielded a number of dinosaur remains, as well as remains of other terrestrial and marine vertebrates. The ornithodirans are represented in the Blue Lias by the early pterosaur Dimorphodon macronyx (Buckland, 1829; Owen, 1859; Barrett et al., 2008) the theropods Sarcosaurus woodi (Andrews, 1921; Ezcurra et al., 2020) and Dracoraptor hanigani (Martill et al., 2016), as well numerous specimens currently regarded as indeterminate members of Theropoda (see, for example, Ezcurra et al., 2020); amongst these indeterminate theropods are two specimens: the distal portion of a femur, the proximal portions of a tibia and fibula, and a partial femur that is missing its distal and proximal articular surfaces (Figure 1, Figure 2, and Figure 3A-C). These partial hindlimbs, comprising the complete and articulated knee joint (Figure 1A-C, Figure 3A-B), and the partial femur (Figure 2, Figure 3C), were originally assigned to the genus Scelidosaurus by Owen (1859), and later included them within the hypodigm of the newly erected species Scelidosaurus harrisonii (1861).
Subsequent studies reassessed the affinities of the specimens and concluded instead that these specimens represented a theropod or several theropods, and not an early-branching ornithischian as originally thought (e.g., Newman, 1968). Carrano and Sampson (2004), who evaluated several early theropods in their study, concluded idented that articulated knee joint at least probably belonged to a basal, indeterminate tetanurine theropod. Naish and Martill (2007) similarly referred all the specimens to Tetanurae in their comprehensive study of British non-avian dinosaur taxa.
 Benson (2010) discussed the affinities of the specimens as he studied another Jurassic (Bajocian) British theropod taxon Magnosaurus nethercombensis. In Benson’s analyses, the articulated femur, tibia, and fibula taken together were recovered as a coelophysoid. Ultimately Benson (2010) concluded that both specimens were indeterminate members of Theropoda. A similar comment is made in passing in the study of Sarcosaurus woodi, another British Jurassic theropod, carried out by Ezcurra et al. (2020), though no clear reasoning is given for this assignment.
Benson (2010) discussed the affinities of the specimens as he studied another Jurassic (Bajocian) British theropod taxon Magnosaurus nethercombensis. In Benson’s analyses, the articulated femur, tibia, and fibula taken together were recovered as a coelophysoid. Ultimately Benson (2010) concluded that both specimens were indeterminate members of Theropoda. A similar comment is made in passing in the study of Sarcosaurus woodi, another British Jurassic theropod, carried out by Ezcurra et al. (2020), though no clear reasoning is given for this assignment.
This study reassesses the theropodan material discussed above, compares it with other, similar taxa, and analyses it using three recent, large anatomical datasets, in order to try and determine if this material does in fact represent a distinct taxon, or is otherwise diagnosable to a generic level. By analysing this material, this study aims to broaden our understanding of early neotheropod evolution and to further develop the picture of some of the earliest known dinosaur populations within Europe.
MATERIALS AND METHODS
Specimens
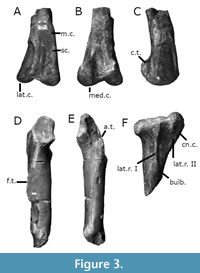 The specimen BMNH 39496 represents a partial but articulated right knee joint comprising the distal end of the right femur and a proximal portion of the right tibia. The femur and tibia were originally found with an associated and articulated right fibula (Figure 1C). However, the whereabouts of this partial fibula is no longer known, and only the articular surfaces upon the proximal end of the tibia where the fibula rested remain (Figure 3F).
The specimen BMNH 39496 represents a partial but articulated right knee joint comprising the distal end of the right femur and a proximal portion of the right tibia. The femur and tibia were originally found with an associated and articulated right fibula (Figure 1C). However, the whereabouts of this partial fibula is no longer known, and only the articular surfaces upon the proximal end of the tibia where the fibula rested remain (Figure 3F).
The cnemial crest is expanded anteriorly and in proximal view is not rounded but hook-shaped; a similar condition is seen in averostran theropods such as Tachiraptor admirabilis (Langer et al., 2014), Piatnitzkysaurus floresi (MACN-CH 895) and Allosaurus fragilis (UMNH VP 7928; Carrano et al., 2012), but unlike the condition in Dilophosaurus wetherilli (e.g., TMM 43636-1) and other, early-occurring ornithischian and sauropodomorph dinosaurs such as Scelidosaurus harrisonii (Norman, 2020). The highest point of this expanded cnemial crest is positioned proximally higher than the other proximal condyles, but only marginally, also similar to the early-branching averostrans Tachiraptor admirabilis and Piatnitzkysaurus floresi (see Bonaparte, 1979). Again, this is in distinction to early diverging ornithischians such as Scelidosaurus harrisonii (Norman, 2020). The lateral face of the later condyle is concave. The lateral and medical condyles are also roughly equally positioned anteroposteriorly. There is a distinct cleft between these two condyles, similar to the condition of many averostrans (e.g., Carrano et al., 2012, fig. 20). The specimen possesses a pronounced fibular crest on the lateral side of the tibia that extends up to the proximal end of the tibia and ends distally as a bulbous expansion (Figure 3F). This same expansion appears in Allosaurus fragilis, Dilophosaurus wetherilli, Megalosaurus bucklandi (OUMNH J.13575) and Piatnitzkysaurus floersi, inter alia, but is not present in Coelophysis bauri (e.g., NMMNH P-29046, Spielmann et al., 2007) or the contemporaneous Sarcosaurus woodi (WARMS G668, WARMS G680; Ezcurra et al., 2020). Parallel to this is a sharp proximodistally oriented crest on the lateral margin of the proximal end of the tibia. This feature is also notably absent in the contemporary taxon Sarcosaurus woodi (WARMS G668) but is present in, for example, Piatnitzkysaurus floresi (MACN-CH 895). However, this unusual second crest does not join to the fibular crest distally to form a forked crest delimiting a proximal fossa, as it does in Piatnitzkysaurus floresi (MACN-CH 895). Posterior to the fibular crest, at around the level of the bulbous expansion, is also small foramen (Figure 3F), also apparently absent in Sarcosaurus woodi (Ezcurra et al., 2020, figs. 10, 11). As noted above, this partial, articular knee joint also previously contained a partial fibula, now lost. Based upon the only available image of this fibula that is available (see Figure 1C, and see Lydekker, 1888), this would have appeared much like the proximally expanded and distally tapered fibulae of many early theropods, e.g., Coelophysis bauri, as well as other early-branching members of Dinosauria, e.g., Heterodontosaurus tucki (Galton, 2014).
Part of the specimen BMNH 39496 is a distal portion of a femur. This distal end of a right femur exhibits an unusual and well-developed proximodistally oriented scar extending along the medial side of the anterior face. There is also a clear medial distal crest present. The medial and lateral condyles extend distally to the same degree, though the medial condyle is anteroposteriorly broader in distal view (Figure 3A-C).
GSM 109560 is a partial left femur (Figure 2, Figure 3D-E). Much of the proximal portion is sadly lost, but the anterior trochanter and fourth trochanter remain. The distal portion of the femur is also missing, meaning little anatomical information can be gleaned of this portion of the bone. What can be observed is a relatively straight and low fourth trochanter (Figure 3E) extruding from approximately the centre of the posterior surface of the femoral shaft. The shaft of the femur itself appears to be relatively robust and straight when viewed anteroposteriorly and mediolaterally; in this respect, the femur of the specimen differs from the condition the contemporaneous Sarcosaurus woodi (WARMS G681; Ezcurra et al., 2020). The proximal portion of the femur in GSM 109560 supports a broadened anterior trochanter; whilst expanded medio-laterally in GSM 109560, this trochanter is not complete aliform, as in, for example, or the more derived Megalosaurus bucklandi, but nor is it quite as narrow and ‘spike-like’ as in the more ‘basal’ Tawa hallae (Nesbitt et al., 2009), Coelophysis bauri (NMMNH P-54620) or Dilophosaurus wetherilli (TMM 43641-1). Crucially, the form of this trochanter differs from that seen in both contemporary theropod taxa Dracoraptor hanigani in which it is wing-like and separated from the shaft by a cleft (Martill et al., 2016), and Sarcosaurus woodi, in which it is barely developed at all and is completely joined to the shaft (Ezcurra et al., 2020). There is again also a distinction with the trochanter as observed in early diverging members of Ornithischia, for example Scelidosaurus harrisonii (Norman, 2020). This third, intermediate condition may well represent a transitional form of this proximal trochanter within Theropoda. GSM 109560 also lacks the trochanteric shelf seen in many other theropods, e.g., Coelophysis bauri (NMMNH P-54620; NMMNH P-29046) and Dilophosaurus wetherilli (TMM 43641-1), and in this way is more akin to taxa such as Allosaurus fragilis and Megalosaurus bucklandi (Benson, 2010).
Institutional Abbreviations
BMNH/NHM UK, Natural History Museum, London, U.K.; GSM, British Geological Survey, U.K.; MACN, Museo Argentino de Ciencias Naturales, Buenos Aires, Argentina; NMMNH, New Mexico Museum of Natural History, New Mexico, USA; OUMNH, Oxford University Museum of Natural History, Oxford, U.K.; TMM, Texas Memorial Museum, Austin, Texas, USA; UMNH, Natural History Museum of Utah, Utah, USA; WARMS, Warwickshire Museum, Warwick, U.K.
SYSTEMATIC PALAENTOLOGY
Clade ARCHOSAURIA Cope, 1869
Superorder DINOSAURIA Owen, 1842
Order THEROPODA Marsh, 1881
Clade AVEROSTRA Paul, 2002
Dornraptor gen. nov.
Type species. Dornraptor normani gen. et sp. nov.
Holotype. BMNH 39496 (Figure 1; Figure 3A-C, F), a partial hind limb. This specimen comprises the distal end of a right femur and a proximal third of a right tibia, originally found in articulation, along with an associated right fibula (now thought to be lost).
Material. GSM 109560 (Figure 2; Figure 3D, E), a partial left femur, still preserving the anterior and fourth trochanters, as well as a trochanteric shelf and a pronounced anterior intermuscular line that is flanked by a foramen.
Basis for referral of GSM 109560. This additional material possesses several anatomical characters that are highly suggestive of a fairly derived position within Neotheropoda, as does the specimen BMNH 39496; both specimens exhibit anatomical features not observed in specimens of the only other known contemporaneous theropodan taxa, i.e., Dracoraptor hanigani and Sarcosaurus woodi. Further, the fact that GSM 109560 was recovered from the same locality within the same stratigraphic unit as BMNH 39496, when considered alongside their similar size and proportions, allows an inference to be drawn that these two specimens likely belong to the same taxon.
Etymology. The generic name, Dornraptor, is derived from Dorn, the shortened version of Dornwaraceasetr, which is the Anglo-Saxon term for the area of England roughly equivalent to the modern County of Dorset, where the material comes from; and raptor, meaning robber or thief, and a commonly employed suffix amongst small-medium sized theropods. Thus, Dornraptor roughly means Dorset robber or thief (Anglo-Saxon, Dorn: “Dorset”; Latin, raptor: ‘one who seizes’). The specific name, normani, honours the notable Cambridge palaeontologist Dr. David B. Norman who has worked extensively on other dinosaur taxa from the Jurassic of the U.K., including Scelidosaurus harrisoni, a taxon to which this material was once erroneously assigned. Dr. Norman has made significant contributions to both the scientific literature on the Dinosauria and to the training of a new generation of palaeontologists from the U.K. and abroad; his impact and legacy cannot be understated.
Age. Hettangian-Sinemurian, Early Jurassic (Cope et al., 1980).
Occurrence. Blue Lias Formation, Lower Lias Group, Charmouth, UK.
Diagnosis. An averostran theropod from the Early Jurassic of Great Britain, Dornraptor normani, can distinguished from all other theropods by its possession of the following unique combination of characteristics (*mark potential autapomorphies): a large proximodistally oriented scar extending along the medial side of the anterior face of the distal end of the femur; possession of a medial distal crest; possession of a well-developed, blade-like anterior trochanter that is separated from the head by a cleft; possession of a low, poorly-developed trochanteric shelf; an anterior surface of the femur that is pierced by a foramen medial to the anterior intermuscular line*; possession of a deep and narrow anterior sulcus between articular condyles of the distal end of the femur; a medial condyle of the femur with an anteroposterior diameter roughly equal to the average mediolateral width of the articular surface of the femur; an antero-posteriorly elongate cnemial crest that reaches further proximally than either the medial or lateral condyles; possession of a pronounced fibular crest on the lateral side of the tibia that extends up to the proximal end of the tibia and ends distally as a bulbous expansion*; presence of a sharp proximodistally oriented crest on the lateral margin of the proximal end of the tibia that runs parallel with the fibular crest, but that does not join to the fibular crest distally to form a forked crest delimiting a proximal fossa; lateral and medial condyles that are roughly similarly placed posteriorly on the distal end of the tibia; possession of a distinct cleft between posterior condyles of the proximal end of the tibia; and possession of a lateral condyle with a posterolateral margin that forms an acute angle when viewed medially*. For further comparisons with other contemporaneous taxa, see above.
Phylogenetic Analyses
The two theropod specimens from the Blue Lias Formation that for the basis of study were first added to the large early dinosaur datasets of Baron et al. (2017a), as modified first by Langer et al. (2017) and then by Baron et al. (2017b). BMNH 39496 and GSM 109560 were then also scored into the more theropod-focused datasets of Zahner and Brinkmann (2019) and of Smith et al. (2010), the latter as further modified by Novas et al. (2015) and then Baron (2024). Specimens BMNH 39496 and GSM 109560 were scored individually and in combination, as a single operational taxonomic unit (OTU) in each dataset. This combination of datasets was chosen specifically because it allows for the broadest possible comparison across the earliest members of Dinosauria, as well as also a more focused set of comparisons with many of the earliest known and best understood members of Theropoda.
With each of the datasets used in this study, trees were searched for using both equal and implied weights implementation of parsimony, using TNT 1.5-beta (Goloboff et al., 2008; Goloboff and Catalano, 2016), through the New Technology search method. Following the protocol of Baron et al. (2017a, b) and Langer et al. (2017), memory was first set at its maximum of 99,999, and trees searched for under equal weights parsimony through a New Technology (Goloboff et al., 2008) search, with ratchet and drift set at their default values and with 100 random additional sequences. A second search was then done, in which trees were searched for using a New Technology Search (Goloboff et al., 2008) using the driven search function, until the best score was hit 100 times. The MPTs produced in this second type of analysis were then subjected to a second round of TBR branch swapping, with a change probability of 33 and 100 additional sequences as the default search settings.
RESULTS AND DISCUSSION
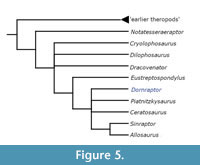
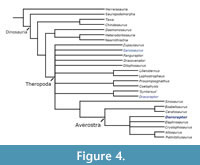 When utilising the modified dataset of Baron et al. (2017a), under equal weights parsimony and with both BMNH 39496 and GSM 109560 combined into a single out, hereafter named as Dornraptor normani this previously difficult to place theropod material is recovered within a small polytomy firmly nested within Neotheropoda (Figure 4). The specimens, when combined, are recovered alongside Elaphrosaurus, Cryolophosaurus and two other pairs of derived theropodan taxa: Allosaurus and Piatnitazkysaurus; and Ceratosaurus and Eoabelisaurus. Adopting the current definition of Averostra (Paul, 2002) as set by Ezcurra and Cuny (2007) - the least inclusive clade including Ceratosaurus and Allosaurus, this analysis concludes that the theropod material now assigned to Dornraptor represents a member of Averostra, sensu stricto. When the specimens are not combined into a single OTU, the majority of Theropoda collapses into a large polytomy. In the analyses that utilised the data of Zahner and Brinkmann (2019), a single combined OTU, Dornraptor normani, is recovered in a polytomy with many definitive averostran theropods, including again Ceratosaurus, Piatnitzkysaurus and Allosaurus (Figure 5). Again, when splitting the specimens apart into two OTUs, the resolution within Theropoda is lost, though unlike in the analyses using the Baron et al. (2017a) data, this approach still placed each specimen within Averostra sensu stricto.
When utilising the modified dataset of Baron et al. (2017a), under equal weights parsimony and with both BMNH 39496 and GSM 109560 combined into a single out, hereafter named as Dornraptor normani this previously difficult to place theropod material is recovered within a small polytomy firmly nested within Neotheropoda (Figure 4). The specimens, when combined, are recovered alongside Elaphrosaurus, Cryolophosaurus and two other pairs of derived theropodan taxa: Allosaurus and Piatnitazkysaurus; and Ceratosaurus and Eoabelisaurus. Adopting the current definition of Averostra (Paul, 2002) as set by Ezcurra and Cuny (2007) - the least inclusive clade including Ceratosaurus and Allosaurus, this analysis concludes that the theropod material now assigned to Dornraptor represents a member of Averostra, sensu stricto. When the specimens are not combined into a single OTU, the majority of Theropoda collapses into a large polytomy. In the analyses that utilised the data of Zahner and Brinkmann (2019), a single combined OTU, Dornraptor normani, is recovered in a polytomy with many definitive averostran theropods, including again Ceratosaurus, Piatnitzkysaurus and Allosaurus (Figure 5). Again, when splitting the specimens apart into two OTUs, the resolution within Theropoda is lost, though unlike in the analyses using the Baron et al. (2017a) data, this approach still placed each specimen within Averostra sensu stricto.
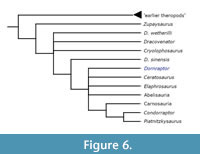
 Finally, using a modified version of the Smith et al. (2010) theropod dataset, the results again suggest that D. normani represents and early-branching averostran theropod (Figure 6). This consensus tree was formed from 53 MPTs each having a length of 1285 steps and positions D. normani within Averostra sensu stricto, in a polytomy with two large clades of derived theropods, such as the Abelisauroids and the Coelurosaurs, as well as Ceratosaurus and Elaphrosaurus.
Finally, using a modified version of the Smith et al. (2010) theropod dataset, the results again suggest that D. normani represents and early-branching averostran theropod (Figure 6). This consensus tree was formed from 53 MPTs each having a length of 1285 steps and positions D. normani within Averostra sensu stricto, in a polytomy with two large clades of derived theropods, such as the Abelisauroids and the Coelurosaurs, as well as Ceratosaurus and Elaphrosaurus.
Both the specimens BMNH 39496 and GSM 109560 possess anatomical characters that are suggestive of a derived position within Neotheropoda and even Averostra. Though these specimens have no anatomical overlap, making a direct comparison between them impossible, their discovery within the same stratigraphic unit, their similar size and proportions, and the fact that they each exhibit a suit of ‘more derived’ theropodan traits that distinguish them from many earlier-branching forms (e.g., from coelophysids and other non-neotheropods), and from contemporaneous taxa, all means that it can be reasonably inferred that these specimens belong to the same taxon. The only other theropods known from the same formation, Dracoraptor hanigani and Sarcosaurus woodi, are both quite distinct from BMNH 39496 and GSM 109560 in terms of the anatomical characters they exhibit (see above).
 Further, the dataset of Baron et al. (2017a) also contained both Dracoraptor hanigani and Sarcosaurus woodi, and the analysis that was carried out using this data clearly recovered the newly created Dornraptor normani in a more deeply nested position within Neotheropoda than either Dracoraptor hanigani or Sarcosaurus woodi (Figure 4). This again suggests that this material represents a more ‘derived’, Averostran theropod than those taxa. Similar results were achieved using two other anatomical datasets (see above). Hence, this study concludes that we can be confident that these specimens do represent a ‘more derived’ theropod than those already named from the Early Jurassic of England.
Further, the dataset of Baron et al. (2017a) also contained both Dracoraptor hanigani and Sarcosaurus woodi, and the analysis that was carried out using this data clearly recovered the newly created Dornraptor normani in a more deeply nested position within Neotheropoda than either Dracoraptor hanigani or Sarcosaurus woodi (Figure 4). This again suggests that this material represents a more ‘derived’, Averostran theropod than those taxa. Similar results were achieved using two other anatomical datasets (see above). Hence, this study concludes that we can be confident that these specimens do represent a ‘more derived’ theropod than those already named from the Early Jurassic of England.
Dornraptor normani also constitutes one of the oldest known theropods from the U.K.; only the early diverging coelophysoid theropod Pendraig milnerae (Spiekman, 2021) is known from older strata. This conclusion therefore adds to our current understanding of early theropod evolution and distribution within the United Kingdom and Europe more broadly and suggests the presence of Averostra in England during the Early Jurassic. This taxon seems to have been a contemporary of other, non-averostran theropods, as well as a numerous other vertebrates, including at least one thyreophoran ornithischian dinosaur (Norman, 2020), and this picture of Early Jurassic terrestrial fauna is consistent with other localities of a similar age globally. We see a similar combination of early ornithischians/possible thyreophoran ornithischians and early branching averostran theropods in the La Quinta formation of Venezuela, for example, i.e., Laquintasaura venezuelae and Tachiraptor admirabilis (Barrett et al., 2014; Langer et al., 2014).
Finally, this study also emphasizes the benefit of constantly reappraising material housed in historic collections. Our broader understanding of non-avian dinosaur diversity and distribution throughout the Mesozoic can be expanded not just with new discoveries in the field, but also with such reappraisal and fresh analyses of older materials (Tennant et al., 2018); hitherto hidden patterns of diversity and disparity within existing collections may continue to be uncovered as more powerful analytic tools and datasets become available and more new taxa named and described for comparison (see for recent examples, Agnolin and Rozadilla 2017; Ezcurra et al., 2020). It is only by constant reappraisal of known material, often in light of new discoveries, that we can ever truly understand the true richness of the global fossil record.
ACKNOWLEDGEMENTS
 The author is extremely grateful to P.M. Barrett and D.B. Norman for their supervision, assistance, guidance, and friendship during his academic progression. The author is also very grateful to both reviewers for taking the time to carefully consider the manuscript in its earlier forms and for the helpful comments that have greatly improved the final version. Megan Williams is also to be thanked wholeheartedly for providing the excellent original artwork used for Figure 7 of the manuscript.
The author is extremely grateful to P.M. Barrett and D.B. Norman for their supervision, assistance, guidance, and friendship during his academic progression. The author is also very grateful to both reviewers for taking the time to carefully consider the manuscript in its earlier forms and for the helpful comments that have greatly improved the final version. Megan Williams is also to be thanked wholeheartedly for providing the excellent original artwork used for Figure 7 of the manuscript.
REFERENCES
Agnolín, F.L. and Rozadilla, S. 2017. Phylogenetic reassessment of Pisanosaurus mertii Casamiquela, 1967, a basal dinosauriform from the Late Triassic of Argentina. Journal of Systematic Palaeontology, 16:853–879.
https://doi.org/10.1080/14772019.2017.1352623
Andrews, C.W. 1921. On some remains of a theropodous dinosaur from the Lower Lias of Barrow-on-Soar. Annals and Magazine of Natural History, 9:570–57.
Baron, M.G. 2024. The effect of character and outgroup choice on the phylogenetic position of the Jurassic dinosaur Chilesaurus diegosaurezi. Palaeoworld, 33:142–151.
https://doi.org/10.1016/j.palwor.2022.12.001
Baron, M.G., Norman, D.B., and Barrett, P.M. 2017a. A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature, 543:501–506.
https://doi.org/10.1038/nature21700
Baron, M.G., Norman, D.B., and Barrett, P.M. 2017b. Baron et al. reply. Nature, 551:E4–E5. Bibcode:2017Natur.551E...4B.
https://doi.org/10.1038/nature24012
Barrett, P.M., Butler, R.J., Edwards, N., and Milner, A. 2008. Pterosaur distribution in time and space: An atlas. Zitteliana Reihe B: Abhandlungen der Bayerischen Staatssammlung fur Palaontologie und Geologie, 28:61–107.
Barrett, P.M., Butler, R.J., Mundil, R., Scheyer, T.M., Irmis, R.B., and Sánchez-Villagra, M.R. 2014. A palaeoequatorial ornithischian and new constraints on early dinosaur diversification. Proceedings of the Royal Society B. Royal Society, 281:20141147.
https://doi.org/10.1098/rspb.2014.1147
Benson, R. 2009. The taxonomy, systematics and evolution of the British theropod dinosaur Megalosaurus. PhD thesis. University of Cambridge, UK.
Benson, R. 2010. The osteology of Magnosaurus nethercombensis (Dinosauria, Theropoda) from the Bajocian (Middle Jurassic) of the United Kingdom and a re-examination of the oldest records of tetanurans. Journal of Systematic Palaeontology, 8:131–146.
https://doi.org/10.1080/14772011003603515
Bonaparte, J.F. 1979. Dinosaurs: a Jurassic assemblage from Patagonia. Science, 205:1377–1379.
https://doi.org/10.1126/science.205.4413.1377
Buckland, W. 1824. Notice on the Megalosaurus, or great fossil lizard of Stonesfield. Transactions of the Geological Society of London, 2:390–396.
https://doi.org/10.1144/transgslb.1.2.390
Carrano, M.T. and Sampson, S.D. 2004. A review of coelophysoids (Dinosauria: Theropoda) from the Early Jurassic of Europe, with comments on the late history of the Coelophysoidea. Neues Jahrbuch fur Geologie und Palaontologie Monatshefte, 2004:537–558.
Carrano, M.T., Benson, R., and Sampson, S.D. 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda). Journal of Systematic Palaeontology, 10:211–300.
https://doi.org/10.1080/14772019.2011.630927
Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Transactions of the American Philosophical Society, 14:1–252.
https://doi.org/10.2307/1005355
Cope, J.C.W., Getty, T.A., Howarth, M.K., Morton, N., and Torrens, H.S. 1980. A correlation of Jurassic rocks in the British Isles. Part One: Introduction and Lower Jurassic. Geological Society of London, Special Report, 14:1–73.
Ezcurra M.D. and Cuny, G. 2007. The coelophysoid Lophostropheus airelensis, gen. nov.: A review of the systematics of “Liliensternus” airelensis from the Triassic-Jurassic outcrops of Normandy (France). Journal of Vertebrate Paleontology, 27:73–86.
https://doi.org/10.1671/0272-4634(2007)27[73:TCLAGN]2.0.CO;2
Ezcurra, M.D., Butler, R.J., Maidment, S.C.R., Sansom, I.J., Meade, L.E., and Radley, J.D. 2020. A revision of the early neotheropod genus Sarcosaurus from the Early Jurassic (Hettangian-Sinemurian) of central England. Zoological Journal of the Linnean Society, 191:113–149.
https://doi.org/10.1093/zoolinnean/zlaa054
Galton, P.M. 2014. Notes on the postcranial anatomy of the heterodontosaurid dinosaur Heterodontosaurus tucki, a basal ornithischian from the Lower Jurassic of South Africa. Revue de Paléobiologie, 1:97–141.
Goloboff, P.A., Farris, J.S., and Nixon, K. 2008. TNT, a free program for phylogenetic analysis. Cladistics, 24:774–786.
https://doi.org/10.1111/j.1096-0031.2008.00217
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics, 32:221–238.
https://doi.org/10.1111/cla.12160
Langer, M.C., Rincón, A.D., Ramezani, J., Solórzano, A., and Rauhut, O.W.M. 2014. New dinosaur (Theropoda, stem -Averostra) from the earliest Jurassic of the La Quinta formation, Venezuelan Andes. Royal Society Open Science, 1:140–184.
https://doi.org/10.1098/rsos.140184
Langer, M.C., Ezcurra, M.D., Rauhut, O.W.M., Benton, M.J., Knoll, F., McPhee, B.W., Novas, F.E., Pol, D., and Brusatte, S.L. 2017. Untangling the dinosaur family tree. Nature, 551:E1–E3.
https://doi.org/10.1038/nature24011
Lydekker, R. 1888. Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History), Cromwell Road, S.W., Part 1. Containing the Orders Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia, and Proterosauria. British Museum of Natural History, London.
Mantell, G.A. 1825. VIII. Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex. By Gideon Mantell, F. L. S. and M. G. S. Fellow of the College of Surgeons, &c. In a letter to Davies Gilbert, Esq. M. P. V. P. R. S. &c. &c. &c. Communicated by D. Gilbert, Esq. Philosophical Transactions of the Royal Society, 115:179–186.
https://doi.org/10.1098/rstl.1825.0010
Martill, D. 2016. The Oldest Jurassic Dinosaur: A basal neotheropod from the Hettangian of Great Britain. PLoS ONE, 11:e0145713.
https://doi.org/10.1371/journal.pone.0145713
Marsh, O.C. 1881. Principal characters of American Jurassic dinosaurs. Part V. The American Journal of Science and Arts, 3:417–423.
Naish, D. and Martill, D. 2007. Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: Basal Dinosauria and Saurischia. Journal of the Geological Society, 164:493–510.
Nesbitt, S.J., Smith, N.D., Irmis, R.B., Turner, A.H., Downs, A., and Norell, M.A. 2009. A complete skeleton of a Late Triassic saurischian and the early evolution of dinosaurs. Science, 326:1530–1533.
https://doi.org/10.1126/science.1180350
Newman, B.H. 1968. The Jurassic dinosaur Scelidosaurus harrisoni Owen. Palaeontology, 11:40–43.
Norman, D.B. 2020. Scelidosaurus harrisonii (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: biology and phylogenetic relationships. Zoological Journal of the Linnean Society, 191:1–86.
https://doi.org/10.1093/zoolinnean/zlaa061
Novas, F.E., Salgado, L., Suárez, M., Agnolín, F.L., Ezcurra, M.D., Chimento, N.S.R., de la Cruz, R., Isasi, M.P., Vargas, A.O., and Rubilar-Rogers, D. 2015. An enigmatic plant-eating theropod from the Late Jurassic period of Chile. Nature, 522:331–334.
https://doi.org/10.1038/nature14307
Owen, R. 1859. Palaeontology. Encyclopaedia Britannica, 17:91–176.
Owen, R. 1861. Monograph of the fossil Reptilia of the Liassic formations. Part I. A monograph of the fossil dinosaur (Scelidosaurus harrisonii Owen) of the Lower Lias. Palaeontolographical Society Monographs, 13:1–14.
Owen, R. 1862. Monographs on the British Fossil Reptilia from the Oolitic Formations. Part second, containing Scelidosaurus harrisonii and Pliosaurus grandis. Palaeontolographical Society Monographs, 1–16.
Paul, G. 2002. Dinosaurs of the Air. The Johns Hopkins University Press, Baltimore and London.
Raven, T.J. and Maidment, S.C.R. 2017. A new phylogeny of Stegosauria (Dinosauria, Ornithischia. Palaeontology, 2017:401–408.
https://doi.org/10.1111/pala.12291
Smith, N.D., Makovicky, P.J., Agnolin, F.L., Ezcurra, M.D., Pais, D.F., and Salisbury, S.W. 2008. A Megaraptor-like theropod (Dinosauria: Tetanurae) in Australia: support for faunal exchange across eastern and western Gondwana in the Mid-Cretaceous. Proceedings of the Royal Society B, 275:2085–2093.
https://doi.org/10.1098/rspb.2008.0504
Spiekman, S.N., Ezcurra, M.D., Butler, R.J., Fraser, N.C., and Maidment, S.C. 2021. Pendraig milnerae, a new small-sized coelophysoid theropod from the Late Triassic of Wales. Royal Society Open Science, 8:210915.
https://doi.org/10.1098/rsos.210915
Spielmann, J., Lucas, S., Rinehart, L., Hunt, A., Heckert, A., and Sullivan, R. 2007. Oldest records of the Late Triassic theropod dinosaur Coelophysis bauri. New Mexico Museum of Natural History and Science, 41:384–401.
Tennant, J.P., Chiarenza, A.A., and Baron, M. 2018. How has our knowledge of dinosaur diversity through geologic time changed through research history? PeerJ, 6:e4417.
https://doi.org/10.7717/peerj.4417
Zahner, M. and Brinkmann, W.A. 2019. Triassic averostran-line theropod from Switzerland and the early evolution of dinosaurs. Nature Ecology and Evolution, 3:1146–1152.
https://doi.org/10.1038/s41559-019-0941-z
