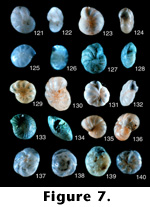
 FIGURE
7.121-140.
FIGURE
7.121-140.Elphidium matsukawauraense Takayanagi (1955, p. 51, figure 27a-27 b).
7.121. Side view, 45x (LD = .24 millimeters).
7.122. Edge view showing aperture, 45x (LD = .24 millimeters).
Photographs of holotype (#67150) in the Tohoku University collections. This specimen is badly etched and could be one of several species. We suspect that it is a distinct species, but need better preserved specimens to verify its species characteristics.
Elphidium sendaiense Takayanagi (1950, p. 26, figure 3).
7.123. Side view, 13.5x (LD = 1.33 millimeters).
7.124. Edge view showing aperture, 13.5x (LD = .89 millimeters).
Photograph of holotype (#66104) in the Tohoku University collections. Similar to E. bartletti, especially in the absence of areal apertures on the last chamber face. However, the specimen is etched, so areal apertures may be obscured.
Elphidium somaense Takayanagi (1955, p. 52, figure 28a-28b).
7.125. Side view, 45x (LD = .24 millimeters).
7.126. Edge view, 45x (LD = .24 millimeters).
Photographs of holotype (#67151) in the Tohoku University collections. This is a very interesting species because there is nothing like it in the Atlantic. It does not appear to belong to the genus Elphidium; perhaps it is Nonion or other allied genera, but it is a distinct species.
Elphidium subcrispum Nakamura (1937, p. 139, pl. 11, figure 8a-8b).
7.127. Side view, 13.5x (LD = .89 millimeters).
7.128. Edge view, 13.5x (LD = .96 millimeters).
Photographs of holotype (no number) in the Tohoku University collections. This species certainly is different from the original E. crispum (Linne 1758) on the basis of a large umbilical boss.
Elphidium subgranulosum Asano (1938a, p. 586, pl. 14 (3), figure 4a-4b).
7.129. Side view, 45x (LD = .47 millimeters).
7.130. Slanted edge view, 45x (LD = .44 millimeters).
Photographs of the holotype (#21418) from the Tohoku University collections. This badly etched specimen belongs with E. poeyanum (d'Orbigny 1839) and is not a distinct species.
Elphidium subincertum Asano (1950a, p. 10, figures 56 and 57).
7.131. Side view, 13.5x (LD = 1.18 millimeters).
7.132. Edge view, 13.5x (LD = 1.18 millimeters).
Photographs of holotype (unnumbered) in the Tohoku University collections. This species is almost certainly the same as the original E. incertum (Williamson 1858), complete with the indistinct sutural pores of the original species.
Elphidium taiwanum Nakamura (1937, p. 139, pl. 11, figure 9a-9b).
7.133. Side view, 13.5x (LD = 1.11 millimeters).
7.134. Edge view, 13.5x (LD = 1.03 millimeters).
Photographs of holotype (no number) in the Tohoku University collections. This large tropical form may actually be a Cellanthus species, but it looks like a distinct species nonetheless.
Elphidium yabei Asano (1938a, p. 589, pl. 14 (3), figures 9a-9b and 10a-10b).
7.135. Side view, 13.5x (LD = 1.63 millimeters).
7.136. Edge view showing aperture, 13.5x (LD = 2.80 millimeters).
Photographs of holotype (#21419) in the Tohoku University collections. This species appears very similar to E. bartletti Cushman (1933), but could be a distinct species.
Epistomaria (Epistomariella) miurensis (Kuwano, 1950, p. 315 and 316, figures 3 and 10).
7.137. Ventral view, 7.5x (LD = 2.00 millimeters).
7.138. Dorsal view, 7.5x (LD = 2.00 millimeters).
7.139. Edge view, 7.5x (LD = 2.27 millimeters).
Photographs of paratype (no number) in the Tokyo National Museum. This species was subsequently called Pseudoeponides japonica by Uchio in Asano (1951c). This appears to be a distinct species and takes precedence over Uchio's name.
Epistomaria (Epistomariella) nakazatoensis Kuwano (1950, p. 316, figures 4 and 11).
7.140. Ventral view, 45x (LD = .38 millimeters).
Photographs of ideotype (no number) in Tokyo National Museum. This species was also placed into Pseudoeponides by Asano (1951c). This species could be a variant within the Ammonia beccarii (Linne 1758) group, but has supplementary apertures on the spiral side similar to the genus Helenina. (See also 8.141.)
Epistomaria (Epistomariella) nakazatoensis Kuwano (continued from 7.140).
8.141. Dorsal view, 45x (LD = .42 millimeters).
Epistomaria yabei Asano (1936a, p. 945, pl. 52 (18), figure 1a-1c).
8.142. Ventral view, 10x (LD = 2.20 millimeters).
8.143. Dorsal view, 10x (LD = 2.20 millimeters).
Photographs of holotype (#21392) in the Tohoku University collections. This is one of the most beautiful photographs taken in this collection. This specimen appears to be a distinct species.
Epistominella takayanagii Iwasa (1955, p. 17, text figure 4a-4c).
8.144. Photograph of a ventral view, 45x (LD = .20 millimeters).
8.145. Dorsal view, 45x (LD = .45 millimeters).
Photographs of holotype (#65504) in the Tohoku University collections. This is an important species in some Pleistocene deposits from the eastern North American continental margin where it sometimes composes up to 90% of a fauna with high abundances (Scott 1987). It is similar to E. sandiegoensis Uchio (1960) from offshore California, but this name takes precedence. However, there is another species illustrated later in this paper (Pseudoparella tamana Kuwano) that is also very similar, but it was very badly preserved in Kuwano's collection. Hasegawa has seen other specimens of this species; it differs from E. takayanagi in having a wider periphery and fewer chambers in the last whorl.
Eponides hatakeyamai Iwasa and Kikuchi (1954, p. 192, text figure 6a-6c).
8.146. Dorsal view, 45x (LD = .38 millimeters).
8.147. Ventral view, 45x (LD = .38 millimeters).
Photographs of holotype (#65521) in the Tohoku University collections. As pointed out in Takayanagi and Hasegawa (1987), this species belongs to the genus Gavelinopsis, but is a distinct species.
Eponides orientalis Asano (1937b, p. 117, pl. 16, figure 8a-8c).
8.148. Ventral view, 13.5x (LD = 1.18 millimeters).
8.149. Dorsal view, 13.5x (LD = 1.18 millimeters).
Photographs of hypotype (#67186) in the Tohoku University collections. The slide is listed as the holotype for E. orientalis (Asano 1951a), but the holotype and the 1951 species are invalid because the 1951 species is a primary junior homonym of the 1937 species and the "holotype" specimen becomes a hypotype. The original holotype is probably IGPS# 21394 from Shiogama Bay in the southern inlet of Matsushima Bay east of Sendai. The hypotype designated in 1951 is from the Tertiary of the Muira Peninsula. This is possibly a distinct species, but the specimen itself is in very bad condition; this is especially evident when it is viewed dorsally and it is difficult to see the suture lines.
Eponides sasai Asano (1938b, p. 94, pl. 10, figures 2 and 3).
8.150. Ventral view, 3x (LD = 5.33 millimeters).
8.151. Dorsal view, 3x (LD = 5.56 millimeters).
Photographs of holotype (#21451) in the Tohoku University collections. This species was placed into the genus Discorbis by Takayanagi and Hasegawa (1987). It appears to be a distinct species.
Eponides subpraecinctus Asano (1951c, p. 12, figures 88-90).
8.152. Ventral view, 13.5x (LD = 1.48 millimeters).
8.153. Dorsal view, 13.5x (LD = 1.48 millimeters).
Photographs of holotype (#67189) in the Tohoku University collections. This species was subsequently placed in the genus Helerolepa by Takayanagi and Hasegawa (1987).
Flintina subglobosa Takayanagi (1953, p. 27, pl. 4, figure 2a-2c).
8.154. Side? view, 13.5x (LD = 1.56 millimeters).
8.155. Apertural view, 13.5x (LD = 1.48 millimeters).
Photographs of holotype (#67135) in the Tohoku University collections. There is a question mark by the side view because we cannot really be sure this is a side view. There are no visible chambers on the exterior, but from the apertural view it is clear that this is not a unilocular form. In the original description of Flintina (from Loeblich and Tappan 1964, 1988), this genus is described as porcelaneous, not agglutinated, so the specimens shown here are clearly not in the correct genus. However, without breaking open the type specimens to determine whether they are planispiral, we cannot determine in what genus the species should be placed.
Globocassidulina bisecta Nomura (1983a, p. 73, pl. 2, figures 2 and 3; pl. 2, figure 3a-c; pl. 14, figures 8-12; pl. 15, figures 1-5).
8.156. Side view showing aperture, 45x (LD = .38 millimeters).
8.157. Apertural view, 45x (LD = .38 millimeters).
Photographs of holotype (#97220) in the Tohoku University collections. This species is similar to G. subglobosa (Brady 1881), but the aperture bisects into two slits, one running perpendicular to the coiling direction and one up against the whorl in the last chamber, which Nomura (1983a) showed.
Globocassidulina crenulata Nomura (1983a, p. 60, pl. 1, figures 9 and 10; pl. 19, figures 7-9 and 11).
8.158. Side view showing aperture, 45x (LD = .33 millimeters).
8.159. Apertural view, 45x (LD = .35 millimeters).
Photographs of holotype (#97191) in the Tohoku University collections. This species is very similar to Cassidulina yabei serrata Matsunaga, described earlier here. As with the serrata, there is a serrated edge and the overall form is similar to the Islandiella species so common in the North Atlantic; we put this species into Matsunaga's species, which takes precedence over this name. Hence, this species becomes Cassidulina (Islandiella?) yabei serrata.
Globocassidulina mucronata Nomura (1983a, p. 63, pl. 1, figures 12 and 13; pl. 13, figures 2-4).
8.160. Side view showing aperture, 45x (LD = .35 millimeters).
Photographs of paratype (#97195) in the Tohoku University collections. This specimen is poorly preserved, but Scott assigns this to G. subglobosa . (See also 9.161.)
Globocassidulina mucronata Nomura (continued from 8.160).
9.161. Apertural view, 45x (LD = .35 millimeters).
Globocassidulina neobrocha Nomura (1983a, p. 67, pl. 1, figure 16a-c; pl. 19, figures 3-6).
9.162. Side view showing aperture, 45x (LD = .49 millimeters).
9.163. Edge view showing aperture, 45x (LD = .49 millimeters).
Photographs of holotype (#97206) in the Tohoku University collections. These specimens appear appear to be a distinct species.
Globocassidulina venustas Nomura (1983a, p. 60, pl. 1, figures 7a-7c and 8; pl. 14, figures 4-7).
9.164. Edge view showing aperture, 45x (LD = .20 millimeters).
9.165. Side view; 45x (LD = .20 millimeters).
Photographs of holotype (#97224) in the Tohoku University collections. This is a distinct species; it has an overall external outline similar to that of Islandiella, but it has an extra external coil not visible in Islandiella.
Gyroidina nipponica Ishizaki exigua Ujiie (1977, p. 99, pl. 17, figure 5).
9.166. Ventral view, 45x (LD = .20 millimeters).
9.167. Dorsal view, 45x (LD = .20 millimeters).
Photographs of holotype (#983) in National Science Museum (Tokyo, Shinjuku branch). This subspecies does not appear to be a true Gyroidina because it lacks any umbilical cavity such as is seen in G. soldani (dOrbigny 1826). Hasegawa placed this species into Gyroidina umbonata (Silvestri 1898), which was originally described as Rotalia soldani dOrbigny var. umbonata Silvestri (1898).
Gyroidina suturalis Ujiie (1977, p. 98, pl. 14, figures 8 and 9).
9.168. Ventral view, 45x (LD = .31 millimeters).
9.169. Dorsal view, 45x (LD = .31 millimeters).
Photographs of holotype (#984) in the National Science Museum (Tokyo, Shinjuku branch). This species is also a junior synonym of G. soldanii.
Hanzawaia hamadaensis Asano (1951a, p. 16, figures 21-23).
9.170. Dorsal view, 37.5x (LD = .61 millimeters).
9.171. Ventral view, 37.5x (LD = .61 millimeters).
Photographs of holotype (#67118) in the Tohoku University collections. Specimen is broken but appears to be a distinct species.
Hanzawaia tagaensis Asano (1949, p. 430, figure 1, nos. 28-30; figure 2, nos. 31 and 32).
9.172. Dorsal view, 13.5x (LD = 1.11 millimeters).
9.173. Ventral view, 13.5x (LD = 1.11 millimeters).
Photographs of holotype (#67047) in the Tohoku University collections. This is a very good specimen and looks like a distinct species.
Haplophragmoides hatai Takayanagi, n. sp.
9.174. Edge view showing aperture, 13.5x (LD = 1.70 millimeters).
9.175. Side view, 13.5x (LD = 1.63 millimeters).Holotype: Specimen number 75020 in the Tohoku University collections.
Type locality: Lat. 43o, 03' 24.6"N, Long. 141o, 54' 39.2"E. Locality no. YH-108, cliff of small tributary of the northern branch of the Anoro River, about 1,150 m south of Hinode Shrine, Hinode, Kuriyama-machi, Yubari-gun, Ishikari Province, Hokkaido.
Type formation: East Takinoue Formation
Type level: Miocene
Description: Test free, medium in size, planispiral, partially evolute, exposing earlier coils, umbilical region depressed, periphery subacute, somewhat lobulate in later chambers; plane of coiling of outer whorl, in adult, somewhat displaced from earlier portion, giving slightly asymmetric appearance to test; chambers slightly inflated, increasing very gradually in size as added, about 12 chambers in last whorl; sutures rather distinct, slightly depressed, straight or slightly curved; wall finely agglutinated with considerable cement, surface rather smoothly finished; aperture at base of apertural face of last formed chamber, but usually indistinct; color usually yellowish brown.
Greatest diameter of holotype is 1.07 mm, least is 0.91 mm; greatest thickness is 0.29 mm. Other specimens range in greatest diameter from 0.28-1.10 mm.
Remarks: This species is similar to Haplophragmoides compressa Leroy (1939) from the Miocene of Sumatra, but differs in possessing rather irregularly coiled whorls with subacute periphery and many more chambers. Specific name is given in honour of Dr. Mitsuo Hata, formerly of the Geological Survey of Japan. This species is unlike any ever seen in the Recent and almost appears in outline as a Cyclammina.
Haplophragmoides nishikizawensis Takayanagi, n. sp.
9.176. Side view, 13.5x (LD = 1.48 millimeters).
9.177. Edge view showing aperture, 13.5x (LD = 1.41 millimeters).Holotype: Specimen number 75021 in the Tohoku University collections.
Type locality: Lat. 43o, 00' 32.2"N, long. 141o, 57' 06.8"E. Location no. YH-132, cliff of a small tributary of the Orochinosawa, about 650 m SW of Nishikizawa Station of the Yubari Railway Line, Yubari City, Ishikari Province.
Type formation: East Takinoue Formation
Type level: Miocene
Description: Test free, medium in size, globose, planispiral, involute, slightly umbilicate, periphery rounded, often distorted in various manners; chambers increasing in size gradually as added; about seven in last whorl; sutures nearly radial, usually not so distinct; wall moderately coarse, agglutinated, but surface rather smoothly finished with cement; aperture a narrow low slit at base of apertural face of last formed chamber; color mostly grayish white.
Remarks: Although this species is variously distorted, it resembles closely Haplophragmoides proboscidiformis Smith (1949) from the early Middle Miocene of Louisiana (USA). However, it differs from the latter species in a globose, larger test without an apertural lip.
At first this species appears similar to Cribrostomoides subglobosus (Sars 1872), but that species is not as deformed looking as this species, and unlike C. subglobosus, this species does not appear to have a cribrate aperture structure, but just a slit.
Haplophragmoides sintikuensis Nakamura (1937, p. 133, pl. 10, figure 2a and 2b).
9.178. Side view, 7.5x (LD = 2.80 millimeters).
9.179. Edge view showing aperture, 7.5x (LD = 2.80 millimeters).
Photographs of holotype (#60853) in the Tohoku University collections. This species was subsequently placed in the genus Reticulophragmium by Huang (1968). This may be a distinct species, but is similar to H. hancocki Cushman and McCulloch (1939) from nearshore areas off New England.
Haplophragmoides taiwanensis Nakamura (1937, p. 134, pl. 10, figure 3a and 3b).
9.180. Side view, 13.5x (LD = 1.85 millimeters).
Photographs of holotype (#60854) in the Tohoku University collections. This coarse-grained specimen is a distinct species. (See also 10.181.)
NOTE: LD (longest dimension).
![]()
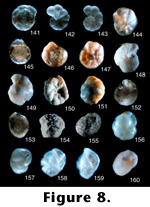 FIGURE
8.141-160.
FIGURE
8.141-160.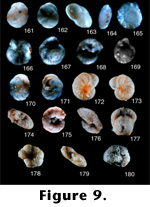 FIGURE
9.161-180.
FIGURE
9.161-180.