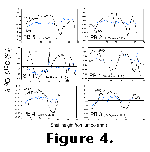
 The
results of isotopic analysis are shown in Figure
4.
The
results of isotopic analysis are shown in Figure
4. The
results of isotopic analysis are shown in Figure
4. ![]() 18O
and
18O
and ![]() 13C
values are plotted on the y-axis (values decreasing upward, as conventional)
against shell height from the origin of growth (except for specimen PB1, where
the early part of the shell had been destroyed; in this specimen height is
expressed relative to the ventral margin). A tabulated version of the full
dataset, together with calculated temperatures, can be obtained as a
downloadable Excel (v.5/95) file.
13C
values are plotted on the y-axis (values decreasing upward, as conventional)
against shell height from the origin of growth (except for specimen PB1, where
the early part of the shell had been destroyed; in this specimen height is
expressed relative to the ventral margin). A tabulated version of the full
dataset, together with calculated temperatures, can be obtained as a
downloadable Excel (v.5/95) file.
All shells analyzed show cyclical patterns of ![]() 18O,
with values mostly ranging between approximately -0.5 and +2.5
18O,
with values mostly ranging between approximately -0.5 and +2.5![]() .
This range, together with the shell-height separation of minima and maxima, is
similar to that observed in modern indigenous A. opercularis
from the southern North Sea (Figure 5),
and, as in these, can be interpreted as the result of seasonal temperature
variation (given equilibrium deposition, demonstrated in modern A.
opercularis, shell isotopic composition is determined only by
temperature and the isotopic composition of ambient water; the latter can be
assumed to be constantsee above).
.
This range, together with the shell-height separation of minima and maxima, is
similar to that observed in modern indigenous A. opercularis
from the southern North Sea (Figure 5),
and, as in these, can be interpreted as the result of seasonal temperature
variation (given equilibrium deposition, demonstrated in modern A.
opercularis, shell isotopic composition is determined only by
temperature and the isotopic composition of ambient water; the latter can be
assumed to be constantsee above). 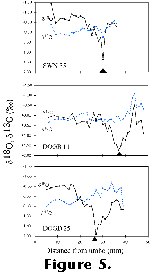 Single-point
large interruptions (in
Single-point
large interruptions (in ![]() 18O
and
18O
and ![]() 13C) to
the cyclical patternat 29 mm from the umbo in PB2, 10 mm in PB4 and 30 mm in
SP1probably constitute a "transient spike" (Krantz
et al. 1988), or analytical artifact, rather than an environmental effect.
Minor peaks and troughs in the profiles are almost certainly related to
environmental influences; similar small fluctuations, greater than levels of
analytical precision, were noted in modern shells and explained by reference to
measured temperature fluctuations that probably relate to movements of fronts
separating water bodies of differing compositions and temperatures (Hickson
et al. 1999).
13C) to
the cyclical patternat 29 mm from the umbo in PB2, 10 mm in PB4 and 30 mm in
SP1probably constitute a "transient spike" (Krantz
et al. 1988), or analytical artifact, rather than an environmental effect.
Minor peaks and troughs in the profiles are almost certainly related to
environmental influences; similar small fluctuations, greater than levels of
analytical precision, were noted in modern shells and explained by reference to
measured temperature fluctuations that probably relate to movements of fronts
separating water bodies of differing compositions and temperatures (Hickson
et al. 1999).
In detail, results from the Holocene shells differ from modern shells. If a ![]() 18O
value of 0.79 (corresponding to a temperature of 13°, roughly halfway between
typical extreme summer and winter temperatures for the southern North Sea; Figure
6) is taken to mark the transition between the warmest ("summer")
and coolest ("winter") halves of the year, then the "summer"
sectors (
18O
value of 0.79 (corresponding to a temperature of 13°, roughly halfway between
typical extreme summer and winter temperatures for the southern North Sea; Figure
6) is taken to mark the transition between the warmest ("summer")
and coolest ("winter") halves of the year, then the "summer"
sectors (![]() 18O
values <0.79; T >13°) are shorter in the Holocene shells than in the
modern shells; the "winter" sectors (
18O
values <0.79; T >13°) are shorter in the Holocene shells than in the
modern shells; the "winter" sectors (![]() 18O
values >0.79; T <13°) are markedly longer in terms of shell height for
the Holocene shells (Figure 4 and Figure
5; Hickson 1997).
As well as exhibiting more extensive winter growth in comparison to modern
shells, Holocene shells generally lack winter growth rings (marking growth
interruptions) and yield larger extreme winter
18O
values >0.79; T <13°) are markedly longer in terms of shell height for
the Holocene shells (Figure 4 and Figure
5; Hickson 1997).
As well as exhibiting more extensive winter growth in comparison to modern
shells, Holocene shells generally lack winter growth rings (marking growth
interruptions) and yield larger extreme winter ![]() 18O
values, corresponding to lower temperatures (Figure
4 and Figure 6).
18O
values, corresponding to lower temperatures (Figure
4 and Figure 6). 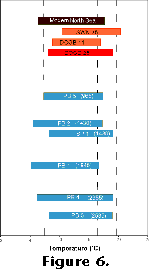 The
temperatures represented by the largest positive
The
temperatures represented by the largest positive ![]() 18O
values of modern shells are several degrees above the coldest temperatures that
the animals experienced (Figure 5 and Figure
6). Given the presence of strong growth rings in modern shells at positions
corresponding to the largest positive
18O
values of modern shells are several degrees above the coldest temperatures that
the animals experienced (Figure 5 and Figure
6). Given the presence of strong growth rings in modern shells at positions
corresponding to the largest positive ![]() 18O
values (Figure 5; Hickson
et al. 1999), this undoubtedly reflects growth cessation during the period
of lowest temperatures. Greater overall winter growth and the general absence of
growth rings suggest, by contrast, that the Holocene shells deposited shell
material throughout the winter and that the (lower) minimum temperatures
registered are representative of the coldest experienced. The slightly less
extensive summer growth of Holocene shells is not matched by the presence of
growth rings; hence, as in modern shells, it can be assumed that growth
continued uninterrupted during the summer and that the highest temperatures
registered are indicative of the warmest temperatures experienced.
18O
values (Figure 5; Hickson
et al. 1999), this undoubtedly reflects growth cessation during the period
of lowest temperatures. Greater overall winter growth and the general absence of
growth rings suggest, by contrast, that the Holocene shells deposited shell
material throughout the winter and that the (lower) minimum temperatures
registered are representative of the coldest experienced. The slightly less
extensive summer growth of Holocene shells is not matched by the presence of
growth rings; hence, as in modern shells, it can be assumed that growth
continued uninterrupted during the summer and that the highest temperatures
registered are indicative of the warmest temperatures experienced.
Some ![]() 13C
profiles show cyclicity (in phase with
13C
profiles show cyclicity (in phase with ![]() 18O),
but this is not represented in every shell investigated, and variation in
18O),
but this is not represented in every shell investigated, and variation in ![]() 13C
(mainly between +0.5 and +1.5
13C
(mainly between +0.5 and +1.5![]() )
is much less than for
)
is much less than for ![]() 18O.
Carbon isotope variations are more difficult to interpret than oxygen; work on
non-biogenic carbonates (Romanek et
al. 1992) has indicated that temperature is not a primary control. Prior
studies of A. opercularis have shown that external rather than
metabolic sources of carbon are the most important (Hickson
et al. 1999); therefore, the likely cause of carbon-isotopic variation in
shells is change in the
18O.
Carbon isotope variations are more difficult to interpret than oxygen; work on
non-biogenic carbonates (Romanek et
al. 1992) has indicated that temperature is not a primary control. Prior
studies of A. opercularis have shown that external rather than
metabolic sources of carbon are the most important (Hickson
et al. 1999); therefore, the likely cause of carbon-isotopic variation in
shells is change in the ![]() 13C
of dissolved inorganic carbon (DIC), which varies in relation to photosynthesis
and decomposition of organic carbon (Arthur
et al. 1983). The patterns observed in A. opercularis
shells (modern as well as Holocene) do not, however, conform to a model (Purton
and Brasier 1997; figure 4a) proposed for a hydrographic setting such as
that of the southern North Sea: shallow, well-mixed water unaffected by
upwelling. Purton and Brasier's model predicts antiphase cyclical variations in
13C
of dissolved inorganic carbon (DIC), which varies in relation to photosynthesis
and decomposition of organic carbon (Arthur
et al. 1983). The patterns observed in A. opercularis
shells (modern as well as Holocene) do not, however, conform to a model (Purton
and Brasier 1997; figure 4a) proposed for a hydrographic setting such as
that of the southern North Sea: shallow, well-mixed water unaffected by
upwelling. Purton and Brasier's model predicts antiphase cyclical variations in ![]() 18O
and
18O
and ![]() 13C, such
that when
13C, such
that when ![]() 18O
values are at their highest (in winter/early spring),
18O
values are at their highest (in winter/early spring), ![]() 13C
values will be at their lowest, due to breakdown of 12C-rich organic
matter; conversely, when
13C
values will be at their lowest, due to breakdown of 12C-rich organic
matter; conversely, when ![]() 18O
values are low due to higher temperatures,
18O
values are low due to higher temperatures, ![]() 13C
values should be at their highest, reflecting uptake of 12C by
phytoplankton. In A. opercularis, however, fluctuations are
either clearly in-phase (e.g., Figure 4e
and 4f) or carbon-isotope variation is not cyclical at all (e.g., Figure
4a). Perhaps this in some way relates to the pattern of variation in
phytoplankton abundance in the southern North Sea, which differs from that of
other temperate shelf seas: at the present time a spring "bloom" does
occur, but the population crash that would normally follow this is not as marked
as it is, for example, in the northern North Sea (Tett
and Walne 1995). In addition, fluctuation in
13C
values should be at their highest, reflecting uptake of 12C by
phytoplankton. In A. opercularis, however, fluctuations are
either clearly in-phase (e.g., Figure 4e
and 4f) or carbon-isotope variation is not cyclical at all (e.g., Figure
4a). Perhaps this in some way relates to the pattern of variation in
phytoplankton abundance in the southern North Sea, which differs from that of
other temperate shelf seas: at the present time a spring "bloom" does
occur, but the population crash that would normally follow this is not as marked
as it is, for example, in the northern North Sea (Tett
and Walne 1995). In addition, fluctuation in ![]() 13C
DIC might conceivably be dampened by decomposition (particularly in
summer) of 12C-rich organic matter supplied by rivers entering the
North Sea or introduced in the form of oil and gas seepages.
13C
DIC might conceivably be dampened by decomposition (particularly in
summer) of 12C-rich organic matter supplied by rivers entering the
North Sea or introduced in the form of oil and gas seepages.
Although there appears to be no pattern (or straightforward explanation) of
short-term variation in ![]() 13C
(i.e., as represented within shells), there is a striking difference in
the range and mean of
13C
(i.e., as represented within shells), there is a striking difference in
the range and mean of ![]() 13C
values from Holocene and modern shells. Most Holocene values are greater
than +0.5
13C
values from Holocene and modern shells. Most Holocene values are greater
than +0.5![]() (mean +0.79; n
= 209); modern values are nearly always less than +0.5
(mean +0.79; n
= 209); modern values are nearly always less than +0.5![]() (mean -0.04; n = 197; sample consisting of all values given by Hickson
1997). The difference in means is statistically significant (F
and t tests;
(mean -0.04; n = 197; sample consisting of all values given by Hickson
1997). The difference in means is statistically significant (F
and t tests; ![]() = 0.05). Conceivably, this difference could be due to higher year-round
productivity in the Holocene (leading to higher
= 0.05). Conceivably, this difference could be due to higher year-round
productivity in the Holocene (leading to higher ![]() 13C
DIC). This would be consistent with evidence (see above) of more extensive
winter growth in the Holocene (winter growth deceleration in modern A.
opercularis may be partly due to reduced food supply rather than low
temperature [Broom and Mason 1978])
and might also explain slightly less extensive summer growth in that
phytoplankton levels above those of the modern North Sea during summer could
cause "clogging" of the gills and slower growth (Chavaud
et al. 1998). However, there is no independent evidence of higher Holocene
productivity and intuitively one would expect modern productivity
to be higher, given the vast present-day supply of nutrients to the North Sea
from domestic and agricultural sources. By contrast, there is voluminous
evidence that, as the result of greater combustion of fossil fuels, modern
atmospheric CO2 levels are higher than in the
pre-industrial Holocene (Friedli et
al. 1986), and that associated reductions in atmospheric
13C
DIC). This would be consistent with evidence (see above) of more extensive
winter growth in the Holocene (winter growth deceleration in modern A.
opercularis may be partly due to reduced food supply rather than low
temperature [Broom and Mason 1978])
and might also explain slightly less extensive summer growth in that
phytoplankton levels above those of the modern North Sea during summer could
cause "clogging" of the gills and slower growth (Chavaud
et al. 1998). However, there is no independent evidence of higher Holocene
productivity and intuitively one would expect modern productivity
to be higher, given the vast present-day supply of nutrients to the North Sea
from domestic and agricultural sources. By contrast, there is voluminous
evidence that, as the result of greater combustion of fossil fuels, modern
atmospheric CO2 levels are higher than in the
pre-industrial Holocene (Friedli et
al. 1986), and that associated reductions in atmospheric ![]() 13C
have been translated through air-sea CO2 exchange into lower modern
values of
13C
have been translated through air-sea CO2 exchange into lower modern
values of ![]() 13C
for DIC and biogenic carbonate (Beveridge
and Shackleton 1994; Böhm
et al. 1996). Like values of
13C
for DIC and biogenic carbonate (Beveridge
and Shackleton 1994; Böhm
et al. 1996). Like values of ![]() 13C
from modern A. opercularis, those from Pliocene
examples are also lower than values from Holocene shells (Johnson
et al. 2000). Given that independent evidence suggests that Pliocene
atmospheric CO2 levels were comparable to present (Kürschner
et al. 1996), it seems reasonable to conclude that the high
13C
from modern A. opercularis, those from Pliocene
examples are also lower than values from Holocene shells (Johnson
et al. 2000). Given that independent evidence suggests that Pliocene
atmospheric CO2 levels were comparable to present (Kürschner
et al. 1996), it seems reasonable to conclude that the high ![]() 13C
values of Holocene A. opercularis shells are a reflection
of low atmospheric CO2.
13C
values of Holocene A. opercularis shells are a reflection
of low atmospheric CO2.
![]()