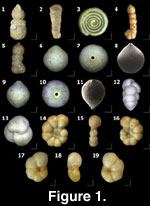
 Figure
1.1 - 1.20.
Figure
1.1 - 1.20.Genus Ammobaculites
Figure
1.1 - 1.20.Ammobaculites jarvisi Cushman
and Renz, 1946
Figure 1.1.
1946 Ammobaculites jarvisi Cushman and Renz: p. 46, pl. 19, fig. 6.
1988 Ammobaculites jarvisi Cushman and Renz; Kaminski et al: p. 188, pl. 4, fig. 4.
1990 Ammobaculites jarvisi Cushman and Renz; Kuhnt and Kaminski: p. 462, pl. 3, fig. d.
1994 Bulbobaculites jarvisi Cushman and Renz; Bolli et al: p. 81, fig. 21.25.
Ammobaculites jarvisi Cushman and Renz, 1946.
Holotype (CC46514) and paratypes (CC46515 and 46516) deposited in the collections of the United States National Museum, Washington, D.C.
Upper Paleocene (P5).
Lizard Springs Formation in Ravine Ampelu, Trinidad.
Test forms a relatively small initial planispire followed by a large, elongate, broad, rectilinear or slightly arched uniserial series, usually with four chambers. Chambers are moderately inflated and separated by depressed sutures. Chamber walls are non-alveolar, well-cemented, finely to coarsely agglutinated, and occasionally include carbonate grains. The primary aperture is a terminal round opening.
Ammobaculites jarvisi Cushman and Renz may agglutinate carbonate material and is characteristic of marly slope environments (Kuhnt and Kaminski 1990). The holotype has an attached test of Ammolagena clavata.
North and South Atlantic, Caribbean.
Common in upper to middle bathyal deposits in Trinidad, the North African paleomargin and the updip wells on the Labrador Margin (Kuhnt and Kaminski, 1990).
Cretaceous (Maastrichtian) - Eocene (Ypresian).
Ammobaculoides
carpathicus Geroch,
1966
Figure 1.2.
1966 Ammobaculoides carpathicus Geroch: p. 479, pl. 13, figs 13-22.
1984 Ammobaculoides carpathicus Geroch; Geroch and Nowak: pl. 1, figs 15-16; pl. 6, figs 8-11.
Ammobaculoides carpathicus Geroch, 1966.
Paratype (USNM 460972) deposited in the collections of the United States National Museum, Washington, D.C.
?Barremian, lower Verovice Shales; in black shale. Also occurs in the upper Cieszyn Shale (Valanginian), and the Grodziszcze Shale (Hauterivian).
Stepina near Strzyzow, Silesia, Poland. Also found at Sulkowice-Ubionka; Lipnik, near Bielsko; and Leszna Gorna, near Goleszow.
Test forms an initial planispire followed by a short biserial series and (finally) elongate uniserial series with numerous chambers. The moderately-inflated chambers are separated by depressed sutures in the uniserial part. This uniserial portion usually forms half to three-quarters of the total length of the test. Uniserial chambers are low and rounded in cross-section, increase gradually in size initially and then become almost constant in size in the (presumptive) adult stage. Wall chambers are medium to coarsely agglutinated with organic cement. The primary aperture is a terminal opening in the uniserial part.
One of the few stratigraphic indicators in Lower Cretaceous sub CCD sediments. Geroch (1966) noted that immersed specimens reveal interiors of chambers, cup-like in the uniserial portion and joined by tubes.
First described from the Polish Carpathians by Geroch (1966). Kaminski et al. (1992) recorded specimens from ODP Site 765 in the Argo Abyssal Plain off northwestern Australia, that were similar in size, outline, and shape of chambers. However, as the wall structure of the Site 765 specimens was not determined, these authors adopted the designation Ammobaculoide cf. carpathicus.
Bathyal to abyssal. Characteristic of sub CCD environments. Lower Cretaceous sub CCD sediments are rare because the CCD was extremely deep in the Tethys and Atlantic Ocean during the Early Cretaceous.
Lower Cretaceous (Valanginian - Aptian) in the Polish Carpathians (Geroch and Nowak 1984).
Ammodiscus latus Grzybowski,
1898
Figure 1.3.
1895 Ammodiscus cf. tenuis Brady; Rzehak: p. 214, pl. 7, fig. 5.
1898 Ammodiscus latus Grzybowski: p. 282, pl. 10, figs 27-28.
1898 Ammodiscus umbonatus Grzybowski: p. 283, pl. 10, figs 29-30.
1948 Lituotuba eocenica Cushman and Renz: p. 7, pl. 1, figs 20-21.
1950 Lituotuba navetensis Cushman and Renz, new name: p. 45.
1981 Ammodiscus latus Grzybowski; Morgiel and Olszewska: p. 8, pl. 1, figs 8-9.
1984 Ammodiscus latus Grzybowski; Geroch and Nowak: pl. 1, figs 12-13; pl. 5, figs 9-12.
1993 Ammodiscus latus Grzybowski; Kaminski and Geroch: p. 254, pl. 5, figs 4a-6.
1994 Ammodiscus latus Grzybowski; Gradstein et al: pl. 9, figs 23-25.
1994 Ammodiscus latus Grzybowski; Bolli et al: p. 68, fig. 18.25.
Ammodiscus latus Grzybowski, 1898.
Depository not given. Lectotype from red clay outcropping in the right bank of the Wislok River at Kroscienlo Nizme, designated and illustrated by Kaminski and Geroch (1993). Lectotype and original syntypic material (No. UJ-132-P, 1/38) deposited in the Grzybowski collection, Jagiellonian University, Krakow, Poland.
Uppermost Eocene; red and grey clay.
Not designated. Localities given: Hanover-Galicia Company wells no. 33 and 34, at Potok; Duniecki well no.1 at Toroszówka; grey clay (rare) and red clay (common) in the bed of the Wislok River at Kroscienlo Nizme; all in the vicinity of Krosno, Poland.
Test forms a robust, thick, evolute, planispiral coil increasing in size very slowly, with a distinct coil suture. In megalospheric individuals, the test consists of two to three whorls with an umbilicus that is depressed on one or both sides. In microspheric individuals, the test comprises up to five whorls, and the umbilicus is usually not depressed. Some specimens display irregular coiling in the last whorl or exhibit a tendency to uncoil. Chamber walls are medium to coarsely agglutinated with organic cement. The primary aperture is simple, at the end of the open tube.
Extremely variable in size.
Worldwide.
Bathyal to abyssal. Common in flysch-type and sub CCD assemblages. The size of tests is probably related to the available carbon flux. For instance, specimens from green organic rich clays of Labrador may exceed 2 mm in diameter, whereas specimens from abyssal ODP Site 647 are smaller, between 600 and 800 microns in diameter (Kuhnt, personal commun., 2001).
Eocene (Bartonian) - Miocene (Messinian). The first common occurrence of A. latus marks the base of the Ammodiscus latus Zone of Geroch and Nowak (1984).
Bulbobaculites
problematicus (Neagu), 1962
Figure 1.4-5.
1962 Ammobaculites agglutinans (d'Orbigny) ssp. problematicus Neagu: p. 61, pl. 2, figs 22-24.
1966 Haplophragmium aequicameratum Huss: p. 68-69.
1974 Haplophragmium lueckei (Cushman and Hedberg); Krasheninnikov: p. 639, pl. 4, figs. 3a-b, 4a, 5a.
1984 Ammobaculites problematicus Neagu; Geroch and Nowak: pl. 1, figs 17-18; pl. 6, fig. 23.
1988 ?Haplophragmium lueckei (Cushman and Hedberg); Moullade et al: p. 363, pl. 3, figs 1-6.
1990 Ammobaculites problematicus Neagu; Kuhnt and Kaminski: p. 465, fig. 5A.
1990 Haplophragmium problematicum (Neagu); Kuhnt: p. 312, pl. 4, figs 3-9.
Ammobaculites agglutinans (d'Orbigny) ssp. problematicus Neagu, 1962.
Depository not given.
Upper Cenomanian-lower Turonian.
Pîrîul Fetii (valea superioara a Buzaului), Carpatii de curbura, red clay.
Test forms a small, initial, streptospiral stage, followed by a uniserial, rectilinear portion of variable length, cylindrical in cross-section. The inflated chambers are separated by markedly-depressed sutures. Chamber walls are finely or coarsely agglutinated with organic cement. The last chamber tapers towards a rounded, terminal aperture, generally possessing a smooth collar.
Bulbobaculites problematicus (Neagu) (including the type specimens in the Neagu collection) displays a wide range of morphologic variability (see Kuhnt and Kaminski 1990, Figures 5, 5A). These authors suggested that the degree of variability was substrate dependent. Specimens from abyssal settings are usually fine-grained, whereas specimens found in turbidites are coarser-grained.
Ammobaculites lueckei Cushman and Hedberg (1941), which is the type species of the Genus Bulbobaculites Maync (1952) is most likely a synonym of Bulbobaculites problematicus (Neagu). However, the only specimen available for taxonomic comparison in the collections of the Smithsonian Institution is the holotype. Although Cushman and Hedberg (1941) reported in the initial description and illustration that the holotype possessed an elliptical aperture at the end of a short neck, a re-examination of the holotype showed that it has a rounded aperture, with a smooth collar. The holotype of A. lueckei has a red colour confirming its fossilization in deep-sea red clay. Ammobaculites lueckei probably represents one growth stage of B. problematicus.
Bulbobaculites problematicus (Neagu) is a very useful maker in the Atlantic, indicating a post Cenomanian/Turonian boundary age.
Worldwide.
Bathyal to abyssal. Common in flysch-type assemblages.
Cretaceous (Cenomanian - Coniacian) in the Polish Carpathians (Geroch and Nowak 1984). However, the first occurrence of Bulbobaculites problematicus (Neagu) in the North Atlantic occurs after the Cenomanian/Turonian boundary (see Kuhnt and Kaminski 1990, Figure 8). In the marginal basins of Zumaya, Gubbio, and the Penibetic the stratigraphic range of Bulbobaculites problematicus (Neagu) extends into the Santonian (see Kuhnt and Kaminski1990, Figure 8).
Caudammina ovulum
gigantea (Geroch),
1960
Figure 1.6-8.
1955 Hormosina ovulum (Grzybowski); Geroch and Gradzinski: pl. 5, fig. 3.
1960 Hormosina ovulum (Grzybowski) var. gigantea Geroch: p. 43, pl. 2, figs 18-19; pl. 11, fig. 6.
1984 Hormosina ovulum gigantea Geroch; Geroch and Nowak: pl. 1, fig. 20; pl. 5, figs 15-16.
1988 Hormosina gigantea Geroch; Moullade et al: p. 365, pl. 2, fig. 11.
1990 Hormosina ovulum gigantea Geroch; Kuhnt and Kaminski: p. 474, pl. 1, figs a-b.
1992 Hormosina ovulum gigantea Geroch; Wightman and Kuhnt: p. 256, pl. 1, fig. 17.
Hormosina ovulum (Grzybowski) var. gigantea Geroch, 1960.
Depository not given.
Not designated. One figured specimen (Figure 18) from the Upper Cretaceous, Senonian, Lower Istebna Beds; other figured specimens (Figure 19) from beds transitional from the Godula Beds (Aptian-Albian to Senonian) - the Lower Istebna Beds. Also occurs in the Upper Istebna Beds (uppermost Cretaceous?-Paleocene) and Ciezkowice Beds (Paleocene).
Not designated. Figured specimen from the Rudnik, near Wadowice (Figure 18), Czarne (Figure 19) and along the Czarna Wiselka stream (Figure 6), all in the Silesian Beskids, Poland.
Test composed of large, spherical chambers arranged in a rectilinear or arcuate series but typically found as single chamber fragments. Chamber size often exceeds 400 microns in width. Chamber wall is multilayered in contrast to stolon connections that are thin, consisting mostly of a single layer often of coarse quartz grains. The difference in wall structure between the chambers and stolons probably accounts for the isolated occurrence of single chamber fragments. The primary aperture is terminal and produced on a tubular neck. Chamber walls are thick, finely agglutinated, and smoothly cemented with organic cement.
Useful stratigraphic marker with a distinct range from the base of the Campanian to the latest Maastrichtian. In the Carpathian flysch, the first occurrence of Caudammina ovulum gigantea marks the base of the early Campanian-Maastrichtian Hormosina ovulum gigantea Zone of Geroch and Nowak (1984).
Caudammina ovulum gigantea exhibits great variability in size, and intergrades with Caudammina ovulum ovulum are common.
Worldwide.
Bathyal and abyssal. Common in flysch-type assemblages.
Cretaceous (Campanian-latest Maastrichtian) in the Polish Carpathian Flysch, according to Geroch and Nowak (1984). Cretaceous (Campanian-Maastrichtian) in the North Atlantic and Labrador (Kuhnt and Kaminski 1990), in the Indian Ocean (Krasheninnikov 1974), and in the Pacific (Wightman and Kuhnt 1992).
Caudammina ovulum ovulum
(Grzybowski), 1896
Figure 1.9-11.
1896 Reophax ovulum Grzybowski: p. 276, pl. 8, figs 19-21.
1973 Hormosina ovulum (Grzybowski); Krasheninnikov: pl. 7, fig. 12.
1988 Hormosina ovulum (Grzybowski); Moullade et al: p. 364, pl. 2, figs 7-9.
1988 Hormosina ovulum ovulum (Grzybowski); Kaminski et al: p. 186, pl. 2, fig. 10.
1990 Hormosina ovulum ovulum (Grzybowski); Kuhnt and Kaminski: p. 474, pl. 1, figs c-e.
1992 Hormosina ovulum ovulum (Grzybowski); Wightman and Kuhnt: p. 257, pl. 2, fig. 2.
1994 Hormosinella ovulum (Grzybowski); Bolli et al: p. 72, figs 19.17-19.
Reophax ovulum Grzybowski, 1896.
Depository not given.
Erroneously reported as Lower Oligocene, lower Tongrian in Ellis and Messina catalogue. The type level is an Upper Cretaceous red clay.
Between 64 and 70 m below the top of an exploratory coal shaft, about 500 m southwest of the courthouse at Wadowice near Krakow, Poland.
Test composed of ovate chambers arranged in a rectilinear or arcuate series, but usually found as single chamber fragments. Chamber wall is multilayered in contrast to stolon connections that are thin, consisting mostly of a single layer often of coarse quartz grains. Differences in wall structure between chambers and stolons probably account for the isolated occurrence of single chamber fragments. The primary aperture is terminal produced on a tubular neck. Chamber walls are thick, finely agglutinated, and smoothly cemented with organic cement.
Caudammina ovulum (Grzybowski) is similar to Caudammina gigantea Geroch in its very smooth surface and narrow connections between the chambers, but has a smaller size (200-400 microns) and more elongated chambers. Caudammina gigantea (Geroch) has a more restricted stratigraphic range (Geroch 1960; Geroch and Nowak 1984; Moullade et al. 1988), and is a useful marker for the end of the Paleocene. Extremely elongated forms show similarities to Hormosina ovuloides (Grzybowski), but can be distinguished from this species by their narrow connections between the chambers and a more symmetrical eggshape.
Worldwide.
Bathyal and abyssal. Common in flysch-type assemblages.
Cretaceous to Paleocene; given as Hauterivian to Paleocene by Geroch and Nowak (1984).
Eggerella bradyi (Cushman),
1911
Figure 1.12-13.
Type species of the genus Eggerella Cushman, 1933.
1884 Verneuilina pygmaea (Egger) (non Egger); Brady: p. 385, pl. 47, figs 4-7.
1911 Verneuilina bradyi Cushman: p. 54, fig. 87, p. 67, fig. 107.
1951 Eggerella bradyi (Cushman); Phleger and Parker: p. 6, pl. 3, figs 1, 2.
1960 Eggerella bradyi (Cushman); Barker: p. 96, pl. 47, figs 4-7.
1994 Eggerella bradyi (Cushman); Jones: p. 51-52, pl. 47, figs 4-7.
1998 Eggerella bradyi (Cushman); Robertson: p. 24, pl. 3, fig. 4.
Verneuilina bradyi Cushman, 1911.
Depository not given.
Type Level:
Recent.
Not designated. Localities mentioned are scattered stations in the North Pacific Ocean, over 1000 fm.
Test nearly conical, oval in cross-section, initially trochospiral, later (in adult portion) reduced to three, and occasionally two chambers per whorl. The highly-inflated chambers increase rapidly in size and are separated by thin, depressed sutures. Chamber walls are finely agglutinated, canaliculate, and smoothly cemented with calcareous cement. The primary aperture is an elongate slit at the base of final chamber, bordered by a narrow lip.
Worldwide.
Typically occurs in deep-water, oligotrophic environments (Altenbach et al. 1999; Kuhnt et al. 1999, 2000, in press). However, it has calcareous cement and is not found in sub CCD sediments. Recorded in water depth of 129-3125 fm at Challenger stations (Jones 1994).
Oligocene (Rupelian) - Recent.
Haplophragmoides
multicamerus Krasheninnikov,
1973
Figure 1.14-16.
1973 Haplophragmoides multicamerus Krasheninnikov: p. 207, pl. 1, figs 1-2.
1974 Haplophragmoides pervagatus Krasheninnikov; Krasheninnikov: p. 635, pl. 1, figs. 4a, b.
1988 Haplophragmoides multicamerus Krasheninnikov; Moullade et al: p. 364, pl. 5, figs 1-2.
1992 Haplophragmoides multicamerus Krasheninnikov; Wightman and Kuhnt, 1992: p. 256, pl. 3, figs 3-5.
Haplophragmoides multicamerus Krasheninnikov, 1973.
Holotype (No 4011/1) deposited in the collections of the Micropaleontological Laboratory, Geological Institute, Academy of Sciences of the USSR, Moscow.
Upper Cretaceous, Santonian-Campanian.
Holotype from Deep Sea Drilling Project (DSDP) Leg 20, Hole 196, Core 2, Section 1, 38-40 cm interval; 6194 m depth on the abyssal floor, west of the Izu-Bonin Trench, Latitude 30° 06.97'N, Longitude 148° 34.49'E.
Test small, planispiral, flattened, evolute, slightly lobulate in outline, oval in cross-section with rounded periphery. Chambers of previous whorls are partly visible through wide, shallow umbilicus. Lateral sides are slightly concave to flat. Eight to 11 inflated, trapeziform chambers in the last whorl, increasing relatively rapidly in height, are separated by straight, depressed sutures. Chamber walls are finely agglutinated with organic cement and show a smooth, glassy finish. The primary aperture is an elongate equatorial slit at the base of the last chamber.
Width of umbilicus varies but inner coil is always partially visible. According to Loeblich and Tappan (1987), this species belongs to the genus Evolutinella.
Useful stratigraphic marker for the Late Cretaceous and bathymetric indicator for abyssal paleoenvironments.
Worldwide.
Recorded from the Pacific Ocean (Krasheninnikov, 1973; Wightman and Kuhnt, 1992), the Indian Ocean (Krasheninnikov 1974), and the North Atlantic (Moullade et al. 1988).
Abyssal, below CCD.
Cretaceous (Santonian-Maastrichtian).
Campanian in the Pacific Ocean (Krasheninnikov 1973). Santonian to Campanian in the Indian Ocean (Krasheninnikov 1974), Santonian to Maastrichtian in the North Atlantic (Moullade et al. 1988).
Haplophragmoides
pervagatus Krasheninnikov,
1973
Figure 1.17-19.
1973 Haplophragmoides pervagatus Krasheninnikov: p. 208, pl. 1, fig. 7.
Haplophragmoides pervagatus Krasheninnikov, 1973.
Holotype (No 4011/7) deposited in the collections of the Micropaleontological Laboratory, Geological Institute, Academy of Sciences of the USSR, Moscow.
Upper Cretaceous, Santonian-Campanian.
Holotype from DSDP Leg 20, Hole 196, Core 2, Section 1, 38-40 cm interval; 6194 m depth on the abyssal floor, west of the Izu-Bonin Trench, Latitude 30° 06.97'N, Longitude 148° 34.49'E.
Test small, planispiral, flattened, slightly evolute, slightly lobulate in outline, oval in cross-section with rounded periphery. Previous chambers of penultimate whorl are partly visible through small, open umbilicus. Lateral sides are slightly concave, almost flat. Approximately six inflated, trapeziform chambers in the last whorl, increasing gradually in size, are separated by straight, depressed sutures. Chamber walls are finely agglutinated with organic cement and show a smooth, glassy finish. The primary aperture is an elongate equatorial slit at the base of the last chamber.
Wightman and Kuhnt (1992) observed transitional forms between Haplophragmoides pervagatus, Haplophragmoides perexplicatus, and Haplophragmoides constrictus, and suggested that these forms may represent end members of a continuous morphologic lineage.
Width of umbilicus varies, but inner coil is always partially visible. According to Loeblich and Tappan (1987), this species belongs to the genus Evolutinella.
Useful stratigraphic marker for the Late Cretaceous and bathymetric indicator for abyssal paleoenvironments.
Worldwide.
Recorded from the Pacific Ocean (Krasheninnikov 1973) as Haplophragmoides perexplicatus from the North Atlantic (Moullade et al. 1988), and as Haplophragmoides ex gr. perexplicatus-constrictus in the Pacific (Wightman and Kuhnt, 1992).
Abyssal, below CCD.
Cretaceous (Santonian-Campanian).
Campanian in the Pacific Ocean (Krasheninnikov 1973), Santonian-Maastrichtian in the Atlantic Ocean (Moullade et al. 1988; Wightman and Kuhnt, 1992)
![]()