DISCUSSION
Coherence Between Data Sets and Validity of
Patterns
The observed decline of diversity near the
beginning and end of each author's study interval is, of course, to be expected,
given the way the data were collected. As the beginning and the end of the study
interval was reached, fewer taxa characteristic of the core of the study
interval are found. An increasing proportion of the taxa present do not belong
to the list of taxa used by the author and are thus not recorded. When
interpreting data of this sort it is therefore necessary to distinguish the bias
towards lower diversity of this 'edge' effect and place most reliance on the
data from within the main study interval of each author. However, in those
intervals where data of different authors overlap, shorter-term trends may still
be of interest. In particular, they can be compared between data sets to check
the consistency of patterns as seen by different authors, and thus provide an
indication as to how accurately the data reflect a primary biologic signal
instead of only artifacts in recording of data. The intervals in which the data
of different authors overlap (early Pliocene, late early/early mid-Miocene, Fig.
3) show a high degree of similarity to each other.
Data from different authors also show a high
degree of coherence in most other estimates of evolutionary change. Although
there are some differences in absolute values, first and last occurrence
patterns and total turnover (Fig. 4), as
well as average longevity values for taxa (Table
5) from different time intervals, all show similar results. Most remarkably,
even the bimodal distribution of taxon longevities is seen in both Lazarus' and
Abelmann's data sets (Fig. 5).
These similarities strongly suggest that the
patterns shown in Figure 3, Figure
4, and Figure 5 are not just
artifacts of different authors' observational methods, but instead reflect
primary patterns of the actual fossil record. Given the well-preserved nature of
the faunas and the lack of obvious bias in the choice of taxa studied, it seems
reasonable to believe that the patterns observed also reflect original patterns
of faunal change in the living faunas. It is therefore valid to compare the
patterns of faunal change to those of environmental change to answer the
question posed at the beginning of this article: are faunal changes in these
planktonic organisms strongly correlated to those of environmental change?
Correlation Between Evolutionary and
Paleoenvironmental Change
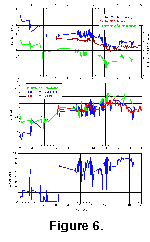 Figure
6 summarizes Kerguelen Plateau Neogene environmental data. Although there
are gaps in these records due both to hiatuses in the sections and occasional
lack of sufficient foraminifera for isotopic analysis (particularly in the late
Neogene), clear patterns are nonetheless apparent. Prior to 15 Ma, oxygen
isotopes were fairly stable and percent sedimentary carbonate values were
uniform and high. Carbon isotope values were increasing, and planktonic values
tended to be heavier than benthic ones. The period between 15 and 13 Ma was one
of major change, with short-term, large-amplitude fluctuations appearing in both
isotope and carbonate records. Oxygen isotopes shifted towards heavier values,
carbon isotopes became more negative, and the offset between benthic and
planktonic values decreased. This new regime persisted to approximately 6.5 Ma,
at which point there was a dramatic drop in carbonate values. There was also
renewed change in both oxygen and carbon isotopes, with both indicators shifting
again in the same direction as in the prior shift at 15-13 Ma. These changes
reflect the well-known Neogene history of the Southern Ocean, which is dominated
by two major events, each of which shifted the system into a new state (Kennett
1982). The changes between 15 and 13 Ma reflect the mid-Miocene increase of
glaciation on the Antarctic continent, together with cooling of both surface and
deep waters in the Southern Ocean, and an increase in biologic productivity (Flower
and Kennett 1993, 1994).
The ca. 6.5-4.5 Ma shift reflects further increases in Southern Ocean
productivity, a shift towards diatom-dominated primary production, and is
associated with a (poorly understood) interval of major glaciation of the
Antarctic continent near the Miocene -Pliocene boundary (Van
Couvering et al. 1976; Kennett
1982).
Figure
6 summarizes Kerguelen Plateau Neogene environmental data. Although there
are gaps in these records due both to hiatuses in the sections and occasional
lack of sufficient foraminifera for isotopic analysis (particularly in the late
Neogene), clear patterns are nonetheless apparent. Prior to 15 Ma, oxygen
isotopes were fairly stable and percent sedimentary carbonate values were
uniform and high. Carbon isotope values were increasing, and planktonic values
tended to be heavier than benthic ones. The period between 15 and 13 Ma was one
of major change, with short-term, large-amplitude fluctuations appearing in both
isotope and carbonate records. Oxygen isotopes shifted towards heavier values,
carbon isotopes became more negative, and the offset between benthic and
planktonic values decreased. This new regime persisted to approximately 6.5 Ma,
at which point there was a dramatic drop in carbonate values. There was also
renewed change in both oxygen and carbon isotopes, with both indicators shifting
again in the same direction as in the prior shift at 15-13 Ma. These changes
reflect the well-known Neogene history of the Southern Ocean, which is dominated
by two major events, each of which shifted the system into a new state (Kennett
1982). The changes between 15 and 13 Ma reflect the mid-Miocene increase of
glaciation on the Antarctic continent, together with cooling of both surface and
deep waters in the Southern Ocean, and an increase in biologic productivity (Flower
and Kennett 1993, 1994).
The ca. 6.5-4.5 Ma shift reflects further increases in Southern Ocean
productivity, a shift towards diatom-dominated primary production, and is
associated with a (poorly understood) interval of major glaciation of the
Antarctic continent near the Miocene -Pliocene boundary (Van
Couvering et al. 1976; Kennett
1982).
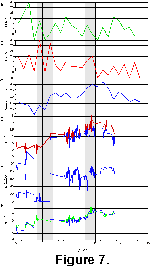 Selected
evolutionary metrics and environmental records are compared to each other in Figure
7. A clear correlation can be seen between the evolutionary (Fig.
7A-C) and environmental (Fig. 7D-F)
data. The overall trends in diversity (Fig.
7C) are correlated to the carbon isotope records (Fig.
7D), particularly that of the planktonic foraminifera, while there is a good
general match between levels of turnover (primarily via stepwise increases in
last occurrences, Fig. 7B) and the three
major phases of Southern Ocean development that are delineated by the two major
intervals of rapid environmental change at 15-13 Ma and near the
Miocene-Pliocene boundary. Faunal diversity thus appears to be inversely
correlated to productivity, as indicated by stable carbon isotope values, and by
the increase in biogenic silica in sediments (indicated by the decrease in
carbonate content in Fig. 7E). This
correspondence is reinforced when other estimates of regional productivity are
considered. Sedimentation rates in Antarctic Neogene pelagic sections for
example tend to reflect water-column productivity, particularly in those later
Neogene biosiliceous sections where carbonate is nearly absent, and thus changes
in carbonate dissolution are not important. Although no comprehensive synthesis
of regional data is available, sedimentation rates in the Antarctic Neogene have
tended to increase from fairly low values in the early Miocene, to moderately
high values in many sections in the late Miocene, reaching a peak in the early
Pliocene before declining into the later Pliocene and Pleistocene (Brewster
1980; Gersonde et al. 1990;
Froelich et al. 1991; Barron
et al. 1991; Harwood et al. 1992).
This pattern, in inverse form, is also reflected in the biodiversity curve (Fig.
7C).
Selected
evolutionary metrics and environmental records are compared to each other in Figure
7. A clear correlation can be seen between the evolutionary (Fig.
7A-C) and environmental (Fig. 7D-F)
data. The overall trends in diversity (Fig.
7C) are correlated to the carbon isotope records (Fig.
7D), particularly that of the planktonic foraminifera, while there is a good
general match between levels of turnover (primarily via stepwise increases in
last occurrences, Fig. 7B) and the three
major phases of Southern Ocean development that are delineated by the two major
intervals of rapid environmental change at 15-13 Ma and near the
Miocene-Pliocene boundary. Faunal diversity thus appears to be inversely
correlated to productivity, as indicated by stable carbon isotope values, and by
the increase in biogenic silica in sediments (indicated by the decrease in
carbonate content in Fig. 7E). This
correspondence is reinforced when other estimates of regional productivity are
considered. Sedimentation rates in Antarctic Neogene pelagic sections for
example tend to reflect water-column productivity, particularly in those later
Neogene biosiliceous sections where carbonate is nearly absent, and thus changes
in carbonate dissolution are not important. Although no comprehensive synthesis
of regional data is available, sedimentation rates in the Antarctic Neogene have
tended to increase from fairly low values in the early Miocene, to moderately
high values in many sections in the late Miocene, reaching a peak in the early
Pliocene before declining into the later Pliocene and Pleistocene (Brewster
1980; Gersonde et al. 1990;
Froelich et al. 1991; Barron
et al. 1991; Harwood et al. 1992).
This pattern, in inverse form, is also reflected in the biodiversity curve (Fig.
7C).
There is no evidence for a pattern of near
uniform, continuous change as predicted by models of evolution driven primarily
by biotic interactions (Stenseth
and Maynard-Smith 1984; Hoffman
and Kitchell 1984). Rates of evolutionary change vary dramatically and are
clearly linked temporally to changes in the environment. Environmental change,
directly or indirectly, thus appears to play a major role in the evolution of
Southern Ocean Neogene radiolarian faunas, with the primary timing and general
magnitude of evolutionary change being driven by external environmental forces.
These results may also be relevant to understanding factors driving the
evolution of Antarctic biotas in general. As noted by Clarke
(1990), temperature change and cold absolute values for temperature, do not
by themselves seem to be sufficient explanations for extinctions of taxa in the
historical development of Antarctic faunas. It may be instead that for much of
the Antarctic biota, and not just for radiolarians, extinction has been caused
by changesprimarily increasesin oceanic productivity, which according to Hallock's
(1987) model, decreased the range of habitats in the 'trophic resource
continuum', and thus led to the loss of biodiversity.
Changes in Evolutionary Rate Characteristics
with Time
One of the more interesting, and quite unexpected,
results of this study are the patterns of longevity data for species. All data
show a similar pattern of bimodal longevities, separated by the relative dearth
of taxa with longevities in the range 10-12 m.y. The subpopulation of long-lived
taxa seems to be found primarily in species that originate in the older part of
the study interval, below 13 Ma, while species of more recent origin are notably
shorter lived. One possible explanation for such a bimodal pattern would be the
existence of two distinct evolutionary sub-groups within the radiolarian
plankton, such as might hypothetically exist between surface and deep-dwelling
faunas. Radiolarians in many parts of the modern ocean include both shallower
dwelling forms and deeper-dwelling species (Casey
et al. 1979; Takahashi 1991;
Casey 1993). The
Southern Ocean, however, is different from lower latitude environments in that
modern radiolarian faunas are most abundant in deeper waters (>150 m to 1,000
m or more) (Abelmann
and Gowing 1996). In the early Neogene, prior to the mid-Miocene climate
shift, surface waters in the Antarctic may have more closely resembled those of
modern cool-temperate regions, with relatively more radiolarian taxa living in
the uppermost layers of the water column. The observed bimodality in
evolutionary rates in the older Neogene data would thus reflect differences in
characteristic evolutionary rates between these two depth environments.
Depth-related differences in evolutionary rates are also known from lower
latitude planktonic foraminifera (Stanley
et al. 1988), where deeper dwelling forms were found to have shorter mean
longevities than the shallower taxa, possibly due to the lower abundances
(population sizes) of deeper dwelling taxa. Extending this idea to the current
study would suggest that the longer lived mode seen in the older Neogene
radiolarian longevity data reflect near-surface dwelling forms. With the loss of
this near-surface water environment in the mid-Miocene, later Neogene
radiolarian faunas became primarily deeper dwellers. The longevity data reflect
this shift in that only shorter lived forms are seen in the late Neogene
longevity histograms. Some support for this interpretation also comes from the
changes in taxonomic abundance between Nassellaria and Spumellaria (see below).
The observed pattern, however, can also be
explained in at least two other ways. First, the bimodality mustat least
partlyreflect the time-windowed nature of these data, plus the substantial
extinction that occurred between 15 and 13 Ma. Many of the taxa lost at the
mid-Miocene event exhibit truncated ranges at the base of the study and thus are
constrained to have longevities <10 m.y. The distribution of the survivors'
longevities suggests that many that do survive then continue to near the second,
Miocene-Pliocene boundary, extinction event, before becoming extinct, thus
creating a second mode of long-lived taxa. However, the same type of explanation
cannot be used to describe the longevity pattern of taxa originating after 13
Ma, as they show a significantly different distribution of longevities, with no
evidence of a large number of long-lived taxa that go through the end-Miocene
event and continue into the Recent. The post mid-Miocene fauna therefore
consists of (on average) significantly shorter ranging species, implying an
increase in evolutionary rates in the region during the Neogene interval
studied.
A second possible alternate explanation is that
not all Southern Ocean taxa are in fact endemic, and in particular, significant
numbers of the post-Miocene taxa appear to be bipolar, with records of
occurrence as well in the Norwegian-Greenland Seas (Bjørklund
1976). Some of the turnover, particularly in the late Neogene, may thus
represent immigration of taxa. This is still of interest but represents a
different biological process for diversity regulation than in situ
evolution (see also discussion in Hoffman
and Kitchell 1984).
It should also be noted that both the overall
rates of change (up to more than 50% species turnover/m.y.) and the average
longevity of species (4.8 m.y.) are comparable to, or exceed, rates that have
been previously reported for marine microplankton groups (ca. 7-16 m.y. for
cladogenetically delimited lineages of planktonic foraminifera (Stanley
et al. 1988); 5.6 and 9.5 m.y. mean longevity for Cretaceous to Recent
keeled and non-keeled species of planktonic foraminifera (including both
cladogenetic and anagenetic evolution, Norris
1991), and ca. 5-10 m.y. for species of planktonic foraminifera,
coccolithophores and radiolarians, based on the survival decay curves given in Hoffman
and Kitchell (1984).
Biological Causes
Although external environmental change appears to
be the primary factor driving evolution of Antarctic Neogene radiolarian faunas,
biology does seem to play an important role. This is suggested by two aspects of
the results. First, diversity and evolutionary turnover metrics appear to
correlate more closely to productivity (carbon isotopes, sedimentation rates),
and the composition of the primary producer flora (carbonate/biogenic silica
ratio), than to temperature itself (oxygen isotopes). This result is similar to
those from the study of the warm water Cenozoic planktonic foraminifera, (e. g.,
Corfield and Shackleton 1988; Lipps
1986; see also discussion in Norris
1991) and Tethyan Mesozoic radiolaria (Erbacher
et al. 1996; Danelian and
Johnson 2001), and lends general support to Hallock's
(1987) trophic model of biodiversity regulation. Second, the shift from a
lower productivity, carbonate phytoplankton ocean to a higher productivity,
biogenic silica phytoplankton ocean is accompanied by a shift in the ecological
nature of the radiolarian fauna (Table 5).
Spumellarians are more common in the older Neogene but are less common in the
later Neogene. Spumellarians are, as a group, more likely to host symbiotic
algae and to live in near-surface waters than are Nassellaria, which tend to be
deeper dwelling, non-symbiotic forms (Casey
et al. 1979; Takahashi 1991;
Anderson 1980, 1993;
Abelmann 1992b). The
shift from spumellarian dominated to nassellarian dominated faunas suggests a
shift from faunas more adapted to lower productivity, silica-rich surface
waters, to faunas living in deeper waters, below a relatively silica poor
surface water layer dominated by highly productive, silica depleting diatoms.
Diatoms are known to be particularly efficient in removing available biogenic
silica in surface waters, and thus are thought to have a potential negative
effect on radiolarian growth (Anderson
1980; Takahashi 1991). On
a much broader scale, Harper and
Knoll (1975) suggested that the Cenozoic evolution of both diatoms and
radiolaria has been influenced by competition for silica. However, it should be
noted that in the modern Southern Ocean, biogenic silica is not a limiting
nutrient - light and water column stability play instead more important roles (Siegfried
et al. 1985). Whether this condition was also true for earlier intervals in
the Neogene is not known.
Conclusions and Suggestions for Future Research
Antarctic Neogene radiolarian faunal occurrence
data drawn from several recent ODP studies have been used to reconstruct the
diversity and evolutionary turnover history of this group, and to compare the
patterns of environmental change. A clear correlation exists between overall
radiolarian diversity and proxies for ocean productivity, such as stable carbon
isotopes, regional sedimentation rates, and abundant biogenic silica in
sediments. Higher rates of inferred productivity characterize the later Neogene
and are associated with lower radiolarian diversity, and an increased dominance
of Nassellarians, a predominantly deeper dwelling group of radiolarians with
fewer photosymbiont taxa.
Overall rates of evolutionary turnover, and
particularly rates of extinction, are not uniform, but are also closely linked
to environmental change, with shifts towards increased turnover and extinction
rates being associated with the shifts in the environment marked by the
mid-Miocene and end Miocene glacial events that affected the Antarctic region.
The average longevity of species also has been
affected by environmental change, with post mid-Miocene species having
significantly shorter stratigraphical ranges than older taxa. This seeming
acceleration of evolution is a phenomenon that has not been reported, or at
least much emphasized in previous studies of warmer water microplankton. Norris
(1991), for example, presents data suggesting that longevity distributions
in Cretaceous to Recent planktonic foraminifera have remained largely unchanged
for the last ca. 100 m.y.
These results together suggest that changes in the
environment, not internal biologic interactions, primarily regulate overall
rates of faunal change in Antarctic Neogene radiolarians. This largely confirms
prior results from study of lower latitude faunas of planktonic foraminifera and
radiolaria, although changing average longevity distributions in Antarctic
Neogene radiolarian faunas suggest that important differences may exist as well,
and that the environmental history of polar and low-latitude regions may differ
in evolutionarily significant ways. Lastly, biology, although not the ultimate
pacesetter of evolutionary change, nonetheless plays an important role in
mediating changes in the physical environment through mixed physical-biologic
mechanisms such as change in marine productivity.
This study is a preliminary analysis, as the
available taxonomic, stratigraphic and biologic data on these faunas are still
very incomplete. Clearly, more complete surveys of the faunas using existing
taxonomy, as well as a more complete taxonomy, would be desirable. So would a
better knowledge of the ecology of the taxa studied, particularly the
relationship between different taxa and marine productivity, including the
distribution of symbionts among species. The relative importance of in situ
evolution vs. immigration from other cold water regions also needs to be
resolved. However, despite these limitations, it appears that useful
evolutionary signals can be extracted even from relatively incomplete data such
as that recorded in normal DSDP/ODP radiolarian biostratigraphic range charts.
Given that few studies of evolution have been carried out with such data sets,
despite their abundance in the published literature, it is hoped that additional
studies may be possible which will yield further insights into patterns and
processes of evolution in the pelagic realm.