INTRODUCTION
An important method for ammonoid identification is the recognition of suture patterns. These patterns mark the union between the septa and the shell wall of the ammonite
phragmocone. Because of the complex structural components of suture patterns, it may be difficult to visually match patterns for species identification. However, the linear nature of the sutures makes them a prime candidate for evaluation using a Geographic Information System (GIS). GIS was developed to enable computers to perform spatial analysis for environmental problems and geological mapping. Using GIS is a novel and innovative way to apply spatial analysis techniques to paleontological problems. A GIS model has been developed that functions as a user-friendly template for matching ammonitic suture patterns for species identification. Unlike other methods, only a fragment of the ammonite that reveals the lateral sutural elements is required -- it is not necessary to break the specimen to see the inner whorls. This model will aid paleontologists and geologists in identifying ammonites for systematics and
biostratigraphy. One advantage of using the GIS sutural template is that GIS software is widely available, and GIS methods are taught in many universities. While developed specifically for species identification, this approach also permits quantification of variation in suture form and, hence, will be a valuable tool for morphometric analyses.
Suture Patterns
Sutures consist of groups of individual elements. These elements transform during the course of phylogeny, and hence offer a method to identify
ammonoids. Two types of sutural terminologies used by paleontologists are morphogenetic and morphographic
(Kullmann and Wiedmann
1970).
Noetling (1905,
1906) first used morphogenetic symbol terminology. This system was improved by
Wedekind (1913,
1916). Noetling and Wedekind's system is based on ontogenetic development that provides information on homologies. Designated symbols are placed on the adult sutural elements, which record the ontogeny of the element, creating a sutural formula.
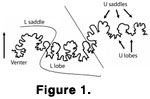
Wedekind's terminology uses five symbols in the order of their phylogenetic appearance. 'E' is the external lobe and external saddle. The 'E' saddle is located on the
venter. The saddle on the venter is cut medially so that one half is on one side of the
ammonoid, and one is on the other. The 'E' lobe is between the 'E' saddle and the 'L' saddle. 'L' is the lateral saddle and lateral lobe. Dorsal to the 'L' lobe is the 'U' saddle and 'U' lobe. 'U' denotes sutures close to the umbilical area of the
ammonoid. 'A' signifies an adventitious lobe. 'I' is the internal saddle and lobe, the saddle and lobe dorsal to the 'U' lobe. 'I' elements are covered by the overlapping shell and so are not usually visible. Juveniles start out with one E, L, U, and I. As the ammonites grow and gain more umbilical lobes, the suture "formula" is denoted as E, L, U2, U, and I. The 'U' subscript gives the order in which the umbilical elements appeared during ontogeny.
Ruzhentsev (1949,
1957) and
Popov (1965) use a modified version of Wedekind's terminology, using different names and symbols for sutural elements.
 Morphographic sutural terminology is a descriptive system based on the position of individual elements.
Schmidt (1921) developed this simpler morphographic method by adding symbols to Wedekind's terminology. This system works well for communication of morphological data.
Morphographic sutural terminology is a descriptive system based on the position of individual elements.
Schmidt (1921) developed this simpler morphographic method by adding symbols to Wedekind's terminology. This system works well for communication of morphological data.
Wedekind's terminology is the oldest practicable morphogenetic system of terminology and is the most popular today. The terminology used in this paper is a modified version of
Wedekind's. For simplicity, only the L and U symbols will be used, because that is all that is necessary for this GIS model of identification
(Figure 1).
The Coilopoceratidae
The Upper Cretaceous Coilopoceratidae Hyatt 1903, which includes the genera
Coilopoceras Hyatt
1903 and Hoplitoides von Koenen 1898, are mostly compressed, involute ammonites that can be either smooth or broadly ribbed, and show slender to somewhat more inflated whorl sections. They have a sharp venter that is narrowly rounded on early whorls and becomes well rounded on the later whorls.
Cobban and Hook (1980) suggested that the
Coilopoceras in the Western Interior of North America were dimorphic with one form being more compressed than the other form. Another dimorphic difference noted by
Cobban and Hook (1980)
is the occurrence of both smooth and ribbed shells. Some specimens even exhibit low, rounded ventrolateral tubercles.
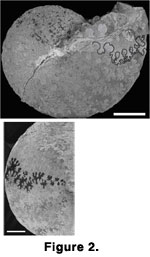 Coilopoceras closely resembles Hoplitoides in appearance and suture pattern, making it difficult to distinguish between the two genera. Both
Coilopoceras and Hoplitoides (Figure
2) have a characteristic suture in which the lateral lobe is broad and deeply bifid, and the dorsal branch of the lateral lobe is in a lower position than the ventral branch.
Cobban and Hook (1980)
stated that
Coilopoceras was derived from Hoplitoides by the total loss of a truncated
venter. Hoplitoides shows a progressive reduction in the extent of venter truncation (constant in each species, but more reduced in younger species), which disappears completely in
Coilopoceras. The difference between Coilopoceras springeri (the oldest
Coilopoceras) and Hoplitoides sandovalensis (the youngest Hoplitoides) is that Coilopoceras springeri completely lacks a truncated venter in the early whorls, whereas Hoplitoides sandovalensis retains a truncated venter in the early whorls.
Coilopoceras closely resembles Hoplitoides in appearance and suture pattern, making it difficult to distinguish between the two genera. Both
Coilopoceras and Hoplitoides (Figure
2) have a characteristic suture in which the lateral lobe is broad and deeply bifid, and the dorsal branch of the lateral lobe is in a lower position than the ventral branch.
Cobban and Hook (1980)
stated that
Coilopoceras was derived from Hoplitoides by the total loss of a truncated
venter. Hoplitoides shows a progressive reduction in the extent of venter truncation (constant in each species, but more reduced in younger species), which disappears completely in
Coilopoceras. The difference between Coilopoceras springeri (the oldest
Coilopoceras) and Hoplitoides sandovalensis (the youngest Hoplitoides) is that Coilopoceras springeri completely lacks a truncated venter in the early whorls, whereas Hoplitoides sandovalensis retains a truncated venter in the early whorls.
The age relationships within the Coilopoceratidae in the Cretaceous Western Interior Seaway of North America are generally sequenced as:
Hoplitoides wohltmanni von Koenen 1897 (the oldest species) followed by Hoplitoides sandovalensis
Cobban and Hook
1980,
Coilopoceras springeri Hyatt 1903,
Coilopoceras colleti Hyatt 1903,
and Coilopoceras inflatum Cobban and Hook
1980 (considered the youngest
Coilopoceras species by Cobban and Hook (1980)).
