DISCUSSION
When rainfall occurs
in this part of Abu Dhabi
it can be extremely violent, resulting in flash floods in gully systems. It might be expected, therefore, that burial of bones would be slow, transport of bones in gullies would be rapid, and that the extremes of weathering would result in the rapid breakdown of bones exposed on the surface. The aim of this monitoring survey was to gain some understanding of the rates of dispersal and weathering of bones in a sub-tropical desert environment. There have been few long-term studies of this nature published, and none at all in this kind of environment, so that even though this study is based on a single skeleton, we feel it offers some insight into taphonomic processes.
The period of the survey was 15 years, from January 1984 to November 1998, and this is the minimum time the bones from the camel skeleton were exposed in the Arabian desert. Total exposure was probably closer to 17 years, since the camel was already largely disarticulated and soft tissues had almost totally decayed. By analogy with other dead animals that we observed in Abu Dhabi, we estimate that it probably died at least two years before the monitoring began.
Weathering
Weathering has been defined by
Behrensmeyer (1978) as destruction of bone by physical and chemical agents operating either on the surface or within the soil. We distinguish between surface weathering, where bone is exposed to variations in heat, moisture and solar radiation, and subsurface weathering, where chemical solution and plant action are more important (Trueman et al. 2004).
Behrensmeyer (1978) considered the physical stresses of heating/cooling and wetting/drying to be the primary causes of surface weathering, and more recent work on chemical and physiological changes in bones exposed to weathering (Trueman et al. 2004) has stressed the heating and cooling effect as instrumental in the breakdown of collagen. For bones buried in soil or in close contact with soil,
Trueman et al. (2004) have additionally proposed a mechanism for replacement of chemical elements in bone by means of water flow whereby water is wicked up from the underside of the bone and evaporated from the exposed side, leading to deposition of authigenic minerals in the pore spaces left open from the collagen decay.
It is clear from the work of
Trueman et al. (2004) that greater precision of environmental variables is needed for a better understanding of the effects of weathering. Their study on mineralization changes in bone was carried out in semi-arid conditions in East Africa, at Amboseli, where rainfall was from 350 to 400 mm per year (Trueman et al. 2004). The rainfall at Jebel Barakah averaged just under 100 m per year for the study period (Figure 2), and temperature varied from 12 to 40º during the year (and sometimes varied by almost as much within a 24 hour period). These climate conditions are probably more extreme than at Amboseli, although comparative data are not available, but it is likely that moisture is less of an issue in the Arabian Desert where rainfall is so much lower. In addition, the sandy substrate does not hold moisture well, and so while it is likely that the effects of temperature are greater at Jebel Barakah than at Amboseli, the effects of mineral replacement by water transport are probably less. We are currently undertaking a programme investigating mineralogical changes in this and other non-tropical environments (Fernandez-Jalvo and Andrews, personal commun., 2005), and the following discussion is restricted to surface modifications of the camel skeleton that were observed in the field and subsequently in the laboratory.
Surface Weathering
Over the period of 15 years (17 assuming an additional two years before observations began), weathering of the bones can be categorized in three modes.
-
For bones exposed on the surface, weathering after eight years (10 years estimated) had barely reached stage 1. After 10 years (12 years estimated), most bones were intermediate between stages 1 and 2, with at least one bone with its most exposed area at stage 3. Little change was seen in the following year, and by 1998, after 15 years exposure (17 years estimated), weathering was well advanced on most bones, with extensive splitting and flaking characteristic of stage 3 surface weathering. This pattern was present mainly on the most exposed parts of the bones, that is those bones that were on the surface all the time and were not buried for any of the time; bones that were stabilized on the surface so that only one side was exposed to weathering; and those parts of the bones that were more vulnerable, such as alveolar bone and processes.
-
Lesser degrees of surface weathering were seen on bones that were subject to movement, although in general one side was more weathered than the other. After 15 years (17 years estimated), stage 2 weathering was the maximum seen, with the less weathered side intermediate between stages 1 and 2. The same degree of surface weathering was also seen on bones that had been buried for part of the time, or had only a shallow covering of sand.
-
Bones more deeply buried had little or no weathering on any surface, so that even after 15 years (17 years estimated) there were two bones that showed no evidence of weathering and several more that were only at stage 1 (Figure 10).
Comparisons can be made with the Amboseli data set of 35 large mammal carcasses on which
Behrensmeyer (1978) based her work. Her study provided a good sample for determining rates of weathering in tropical environments, and the one individual sampled here, albeit for a longer period, is not sufficient to establish a sound time scale for weathering outside the tropics. What can be inferred from this one individual are as follows:
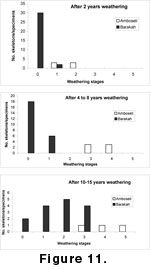 After 2-4 years, most bones visible on the camel skeleton were not weathered, and only two bones had incipient stage 1 weathering, whereas the Amboseli skeletons were all at stages 1 and 2 (see
Figure 11);
After 2-4 years, most bones visible on the camel skeleton were not weathered, and only two bones had incipient stage 1 weathering, whereas the Amboseli skeletons were all at stages 1 and 2 (see
Figure 11);
From years 4 to 8, the bones from the camel skeleton were still mainly unweathered, with less than one quarter at stage 1, whereas the majority of skeletons from Amboseli had reached stages 3-4 (Figure 11);
From years 10 to 15, the majority of bone from the camel skeleton were at stage 2, with a range from 0 to 3, while the Amboseli skeletons had all reached stage 3, with a range from 3 to 5 (Figure 11).
The earliest that any single bone from the camel skeleton had reached stage 3 was by year 12, and no greater degree of weathering was observed after 17 years. In the Amboseli sample, the first skeletons had reached stage 3 by years 4-5, stage 4 by years 6-7, and stage 5 by years 10-15 (Behrensmeyer 1978). It is evident from this data that the climate conditions in Abu Dhabi promote far less rapid weathering than those at Amboseli, but what are these climatic differences? The daily temperature fluctuations in the desert conditions of Abu Dhabi are likely to be greater than at Amboseli, indicating temperature is not a significant factor. If high daily temperature variation was a factor in weathering, the degree and rate of weathering should be more rapid in the desert environment than in tropical savanna, but this is clearly not the case. Seasonal moisture variation would appear to be lower at Jebel Barakah, for rainfall is sporadic and restricted to less than half the year at Jebel Barakah, and daily moisture variation is also less, for there is little dew formed on sandy substrates. Solar radiation is also less at Jebel Barakah, for although the apparent radiation is high in the desert, which makes life quite unpleasant in the absence of any shade, Jebel Barakah is at 24º latitude compared with Amboseli at 2.5º latitude. It would appear likely that the combination of less solar radiation and moisture variation are two of the factors that result in slower weathering in Abu Dhabi.
Subsurface Weathering
Subsurface weathering was readily distinguished from surface weathering. The bones from the camel skeleton that had been underground for any period had light staining and root marks, a phenomenon common to many environments (Andrews and Armour-Chelu 1998). Bones more deeply buried were more deeply stained and had extensive root etching, with roots penetrating below the surface and splitting the bone and spalling off surface bone. Both the splitting and the flaking are deeper than is seen in stages 1-3 of surface weathering, and they are more localised while at the same time exterior bone on other parts of the bone may be little altered. Unfortunately there are too few data on which to set up objective criteria for identifying stages of subsurface weathering, although this is obviously important.
Bone Dispersal
When first observed, the camel skeleton was dispersed over 18 m along the shallow sloping gully with a lateral spread of just under 3 m (Figure 3). This was virtually unchanged after eight years, with an apparently contracted range of dispersal probably as a result of burial of some of the bones (Figure 4). In 1994, 10 years after the skeleton was first monitored, there were several parts of the skeleton 21 to 24 m down the gully (Figure 6), both ribs and limb bones, and these were complete and showed no evidence of carnivore action or rounding. The bones were in or near the line of the channel cutting through the bottom of the gully, and it is most likely that they were transported down the gully by water flow, although there is no direct evidence of this. There was some movement of bones 2-3 m up the gully, and this seems most likely to be due to scavengers or trampling. Four years later, when the monitoring of the camel skeleton finished, there were bones 45 m down the gully (Figure 9). Throughout most of this period, however, the central cluster of bones close to where the camel died remained within the original dispersal area of 14 m and were not transported. This cluster consisted mainly of ribs and vertebrae, which according to experimental work on bone transport are the bones most likely to be moved by water transport (Voorhies 1969,
Lyman 1994).
Rain in Abu Dhabi can be torrential, and flash floods in gully systems are commonplace. There were several periods of heavy rain during the monitoring period, and clearly this had the effect of transporting some of the bones down the gully, but many of the skeletal elements that were moved the furthest (maxilla, astragalus and radioulna), were the ones that in experimental work moved least (Voorhies 1969). The bones remaining at the death site, by contrast, were the ones that are most susceptible to water transport.
Bone Orientations
Orientations of the bones were influenced by their position on the surface, which in turn would be related to surface run-off of water. The bones on the north side of the gully aligned to the south and west, at 221º to 280º, following the slope of the side of the gully towards the channel. The data have been provided in
Figure 3,
Figure 4,
Figure 6, and
Figure 9, but they do not provide evidence independent of topographical slope.
Carnivore Action
 There was surprisingly little evidence of carnivore activity on the camel bones. It was assumed that skeletal elements missing from the skeleton had been removed by scavengers, which in northwestern Abu Dhabi would be most likely to be jackals and foxes, hyaenas being absent from the area now (Harrison and Bates 1990). Chewing was confined to broken edges and near the ends of bone processes and vertebral spines (Figure 10), and the size distribution of pits on diaphyseal surfaces (category a of
Andrews and Fernandez-Jalvo 1997) shows a range overlapping but exceeding that of a larger sample of foxes from southern England (Figure 12). This is consistent with the probable identity of the scavengers being jackals and foxes, but it does not exclude the possibility of larger predators being involved. We have observed at other sites that carnivores of all sizes can make small diameter chewing marks on bones, but small carnivores cannot make large marks, so that it is not so much the range of sizes of marks that distinguish between carnivore chew marks as the greatest size of the marks. The size of the largest carnivore chewing marks on the Jebel Barakah camel skeleton therefore indicate a predator larger than a fox, but unfortunately there is little information on the size of jackal chewing marks. No rodent chewing was observed.
There was surprisingly little evidence of carnivore activity on the camel bones. It was assumed that skeletal elements missing from the skeleton had been removed by scavengers, which in northwestern Abu Dhabi would be most likely to be jackals and foxes, hyaenas being absent from the area now (Harrison and Bates 1990). Chewing was confined to broken edges and near the ends of bone processes and vertebral spines (Figure 10), and the size distribution of pits on diaphyseal surfaces (category a of
Andrews and Fernandez-Jalvo 1997) shows a range overlapping but exceeding that of a larger sample of foxes from southern England (Figure 12). This is consistent with the probable identity of the scavengers being jackals and foxes, but it does not exclude the possibility of larger predators being involved. We have observed at other sites that carnivores of all sizes can make small diameter chewing marks on bones, but small carnivores cannot make large marks, so that it is not so much the range of sizes of marks that distinguish between carnivore chew marks as the greatest size of the marks. The size of the largest carnivore chewing marks on the Jebel Barakah camel skeleton therefore indicate a predator larger than a fox, but unfortunately there is little information on the size of jackal chewing marks. No rodent chewing was observed.
Trampling
Many of the camel bones were unbroken, even after 15 to17 years of exposure. Breakage of vertebral lateral processes and spines was common, most ribs were broken, and the pelvis was broken into two pieces although still complete. This is a type of breakage that even a large carnivore would find difficult, and since the bone was minimally transported it is most likely that the breaks were the result of trampling by a large mammal, most probably other camels, which graze over Jebel Barakah.
