SYSTEMATIC PALEONTOLOGY
Order
Artiodactyla
Suborder
Ruminantia
Infraorder Tragulina
Lophiomerycidae Janis, 1987
Genus
and Species Indeterminate
Material
Locality Z 108: Z 760, fragment of a left P3; Z
761, fragment of a very worn upper right molar; Z 2277, fragment of a right p3;
Z 2278, left mandible with m1-m2; Z 2299, right astragalus.
Locality and Age
Known from only one Zinda Pir site, Locality Z 108
in the lower part of the Chitarwata Formation. Late Oligocene and approximately
25 Ma (Lindsay et al. this issue).
Description
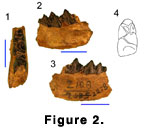 The mandible, Z 2278 (Figure 2.1-2.4) is the only
identifiable specimen. It has the broken posterior root of dp4, intact crown of
m1, and anterior half of a newly erupted m2. Identification of the position of
the teeth is based on the shape of the jaw margin posterior to the m2.
The mandible, Z 2278 (Figure 2.1-2.4) is the only
identifiable specimen. It has the broken posterior root of dp4, intact crown of
m1, and anterior half of a newly erupted m2. Identification of the position of
the teeth is based on the shape of the jaw margin posterior to the m2.
The lower molars are brachyodont, with smooth
enamel, distinctly selenodont protoconid and hypoconid, and a cuspidate
metaconid. On the m2 the unworn protoconid is much taller than the metaconid,
but the slightly worn hypoconid of the m1 appears to have originally been about
the same height as the entoconid. The trigonid is distinctly narrower than the
talonid (trigonid: talonid = 0.89), giving the molars a slightly trapezoidal
aspect in occlusal view. The anterior face of the metaconid is rounded and lacks
any hint of a premetacristid. The antero-lingual face of the protoconid is also
rounded, with the preprotocristid sharply differentiated. After first descending
at a steep angle, the preprotocristid turns abruptly lingually and extends
around the anterior of the tooth as a low crest (“paralophid”), reaching the
lingual margin and having a barely discernable twinned cuspule at its anterior.
The large anterior fossette has a broad lingual opening. The entoconid is
slightly anterior to the hypoconid, and its posterior face is rounded, without a
postentocristid. The antero-lingual face of the entoconid is flat, without an
entoconid groove (“Zhailimeryx-fold”). The hypoconid has a broad and
shallow vertical groove on its posterior face. The posthypocristid extends to
the lingual margin of the tooth, leaving a narrow opening to the posterior
fosset, but without forming a distal tubercle (variously interpreted in closely
related forms as an entostylid or hypoconulid).
There is a distinct “M” shaped structure (“Dorcatherium‑fold”
in the broad sense) on the posterior of the trigonid of both the m1 and m2
(Figure 2.4). The metaconid leg of the “M” (“Dorcatherium‑fold” in the
sense of Janis 1987) is formed from a deep cleft with the resulting lingual fold
reflected labially to form a thickened pillar-like structure. The cleft starts
at the unworn tip of the metaconid and closes before reaching the base of the
cusp. The labial leg on the protoconid (the “Tragulus-fold”) is formed
from an equally deep groove with a reflected and thickened labial margin forming
a ridge connecting to the prehypocristid. The longer postprotocristid and
shorter postmetacristid (the two cross connections of the “M”) are
subhorizontal, coming together at a very shallow angle and joining ventrally
with a vertical crest that connects with the preentocristid. The
postprotocristid, postmetacristid, and vertical crest together form a “Y” shaped
structure on the posterior of the trigonid, with the vertical stem being as long
as the diverging arms. At the base of this structure where it contacts the
preentocristid there is a very small vertical fold in the entoflexid and another
on the labial side making a connection to the “Tragulus‑fold”‑prehypocristid
complex. Finally, there is a strong anterior basal cingulum that arises at the
base of the preprotocristid and then passes around the front of the tooth to
form a lingual tubercle, a very strong and high posterior basal cingulum that
extends labially, and a low bulbous “ectostylid” that is probably a vestige of a
labial cingulum.
The upper molar (Z 761) is very worn and possibly
is an m1. It preserves a faint lingual cingulum, extending around the protocone.
The two premolars are fragments of the anterior of a P3 (Z 760) and posterior of
a p3 (Z 2277). The three specimens preserve little by way of distinctive
morphology and are assigned to this taxon on the basis of size.
Comparisons
This taxon differs from both extant and extinct
tragulids (including Dorcabune
Pilgrim 1910) in the development of the preprotocristid and absence of the premetacristid, as well as in the length of
the common stem of the postmeta- and postprotocristid complex and the closing of
the “Dorcatherium‑fold” before it reaches the base of the metaconid.
Archaeotragulus krabiensis
Métais,
Chaimanee, Jaeger, and Ducrocq 2001, was described as a tragulid, but shares
some similarities with the Zinda Pir mandible not otherwise seen in tragulids.
These principally involve the formation of the trigonid, in that the
preprotocristid of Archaeotragulus krabiensis is developed as a
substantial antero-lingually directed loph that, in combination with the absence
of a premetacristid, leaves the trigonid open lingually. In addition, the
postprotocristid and postmetacristid join at a shallow angle to form a short
vertical stem that passes down the rear of the trigonid to connect to the
preentocristid. There are, however, differences that in our opinion preclude
assigning the Zinda Pir specimen to Archaeotragulus. By comparison to the
Zinda Pir mandible, the lingual cusps of Archaeotragulus krabiensis are
more bulbous, and the unworn trigonid cusps are more equal in height; the
preprotocristid is more lingually directed, and the anterior fosset is less
open; the “Dorcatherium-fold” extends to the base of the metaconid and
the postprotocristid and postmetacristid join at a more acute, steeper angle
with a shorter vertical stem; and there is no sign of a “Zhailimeryx-fold”
on the entoconid nor a distal swelling on the posthypoconid.
The absence of the postentocristid and
premetacristid (producing a “figure-8” morphology), together with strong
anterior and posterior cingula and a wide, lingually open anterior fossette are
characters of taxa placed in the Lophiomerycidae by
Janis (1987) and others (Guo
et al. 2000; Métais et al. 2001). To this character list we would add 1) the “Y”
shaped configuration of the postprotocristid and postmetacristid, with a long
common stem connecting to the preentocristid; 2) the relative heights of the
protoconid and metaconid and the relative lengths of the postprotocristid and
postmetacristid; and 3) the distinctive form of the “Dorcatherium‑fold,”
which is confined to the tip of the metaconid cusp and does not extend to its
base. The presence of these characters in the lower molars of the Zinda Pir form
indicate it is a lophiomerycid.
Of currently recognized lophiomerycids,
Zhailimeryx Guo, Dawson and
Beard 2000 and Krabimeryx
Métais,
Chaimanee, Jaeger, and Ducrocq 2001 are similar to the Zinda Pir specimen in
possession of the typical lophiomerycid features, but differ in many details.
Most notably, differences include the presence of a well defined “Zhailimeryx-fold”
on the anterior face of the entoconid, the apparent absence of a well defined “Tragulus-fold”
connected to the prehypocristid, and the weakness or absence of a bulbous
ectostylid. In addition, in neither of these two taxa does the preprotocristid
extend as far lingually as in the Zinda Pir tooth, although in Zhailimeryx
there is an anterior cusp described by
Guo et al. (2000) as a “rudimentary paraconid” that might correspond to the “twinned cuspule” observed on Z 2278.
Both Lophiomeryx
Pomel 1853 and
Iberomeryx Gabunia 1964 (including Cryptomeryx
Schlosser, 1886) share
the characteristic lophimerycid trigonid features, and both lack the “Zhailimeryx-fold”
on the entoconid, a similarity shared with Z 2278. Nevertheless, Iberomeryx
and especially Lophiomeryx differ substantially from the Zinda Pir
specimen. European and perhaps Asian species of Lophiomeryx are more
hypsodont and lack any vestiges of a “Tragulus-fold” or of basal
tubercles on the labial side of the tooth, while having a more open posterior
fossette, a distinct cuspule terminating the posthypocristid, and an incomplete
postentocristid that extends from the unworn tip of the entoconid partway down
the cusp, leaving the entoconid base rounded. In addition, in Lophiomeryx
the antero-lingually directed prehypocristid joins the
preentocristid‑postmetacristid complex, rather than being more medially
terminated.
Species of Iberomeryx have bulbous basal
tubercles on the labial side of the tooth (interpreted as ectosylids, but
possibly a labial cingulum remnant), a weak “Tragulus-fold” variably
developed on anterior molars, and, also variably, the prehypocristid connected
directly to the trigonid rather than to the preentocristid-postmetacristid
complex (Nanda and Sahni 1990;
Métais et al. 2001). These are similarities
shared with the Zinda Pir mandible and suggest Iberomeryx is the mostly
closely related of known taxa. The Zinda Pir specimen, however, differs from the
better-known species of Iberomeryx such as I. minus and I.
parvus in the strength of the “Tragulus-fold,” which on both its m1
and m2 is very well formed with a thickened labial margin contacting the
anterior end of the prehypocristid. The Zinda Pir mandible also has the terminal
end of the prehypocristid more lingually directed, and there is no trace of a
cuspule on the lingual termination of the posthypocristid.
The enigmatic Iberomeryx savagei
Nanda and Sahni 1990 is known from a few specimens of Late Oligocene age from the Kargil
Formation of northern India. While too small to be the same species, it appears
to also have had a well-formed “Tragulus-fold” contacting the
prehypocristid and a lingually extended preprotocristid.
Discussion
The Tragulina are regarded by most current workers
as a paraphyletic assemblage, and the lophiomerycids may also be paraphyletic,
with some genera having closer relationships to the Tragulidae (Métais et al. 2001). As currently recognized, the family contains Middle Eocene to Late
Oligocene species.
Métais et al.
(2001) suggested the Kargil species
represented a new genus, and it is likely the Zinda Pir specimens are a closely
related species. Forster-Cooper (1915) based Gelocus indicus from Dera
Bugti on a single upper molar. His specimen, which is taken to be a third molar,
has a strong lingual cingulum and very small metaconule. It is of appropriate
size for an upper molar matching Z 2278 and might be the same species.
The “M” structure (or “Tragulus-fold”) is
thought to be a uniquely derived character of tragulids (Métais et al. 2001) or tragulids and leptomerycids (Geraads et al. 1987). The clear presence of the
structure in combination with a lingually extended preprotocristid and absence
of a premetacristid suggests it may be a primitive feature shared by both early
tragulids and lophiomerycids.
