SYSTEMATIC
PALEONTOLOGY
(continued)
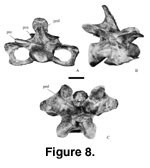 Cervical Vertebrae.
Presacral vertebrae are composed of highly
pneumatized camellate bone (Wedel 2003). The centra are strongly opisthocoelous
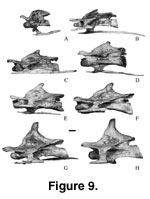
(sensu Romer 1956) with the anterior convexity centrally placed on the centrum. Eighteen cervical vertebrae (Figure 8,
Figure 9,
Table 2), including nine
isolated vertebrae and two articulated sets (Mal-278, three vertebrae; Mal-280,
six vertebrae), of at least two individuals, are attributed to Malawisaurus.
The atlas-axis complex is not represented. Mal-180 (Jacobs et al. 1996, figure
5a), Mal-243, Mal-245, the Mal-278 set, the first four of the Mal-280 set, and
Mal-301 are fairly well preserved while Mal-187-1, Mal-193-1, Mal-244, Mal-246,
the last two of the Mal-280 set, and Mal-291 (= 89-78,
Jacobs et al. 1993,
figure 1e) are poorly preserved.
Cervical Vertebrae.
Presacral vertebrae are composed of highly
pneumatized camellate bone (Wedel 2003). The centra are strongly opisthocoelous
(sensu Romer 1956) with the anterior convexity centrally placed on the centrum. Eighteen cervical vertebrae (Figure 8,
Figure 9,
Table 2), including nine
isolated vertebrae and two articulated sets (Mal-278, three vertebrae; Mal-280,
six vertebrae), of at least two individuals, are attributed to Malawisaurus.
The atlas-axis complex is not represented. Mal-180 (Jacobs et al. 1996, figure
5a), Mal-243, Mal-245, the Mal-278 set, the first four of the Mal-280 set, and
Mal-301 are fairly well preserved while Mal-187-1, Mal-193-1, Mal-244, Mal-246,
the last two of the Mal-280 set, and Mal-291 (= 89-78,
Jacobs et al. 1993,
figure 1e) are poorly preserved.
 In sauropod cervical
vertebrae, the interprezygapophyseal distance, development of laminae and fossae
on the lateral surface of the neural arch, and the length of the diapophyses
increase posteriorly (Osborn and Mook 1921;
Gilmore 1936;
Powell 1986;
McIntosh
et al. 1996a,
1996b). In Haplocanthosaurus (Hatcher 1903), the penultimate
cervical vertebra is wider than high and the last cervical vertebra is shorter
than the preceding cervical vertebra. In Camarasaurus (Osborn and Mook
1921) and Apatosaurus (Gilmore 1936), the lengths of distal cervical
vertebrae are less than the lengths of medial cervical vertebrae. Among titanosaurians, articulated cervical vertebrae are known in “Titanosauridae
indet. DGM Series A” from Brazil, (12 cervical vertebrae, except the atlas,
preserved in articulation with three proximal dorsal vertebrae;
Powell 1986,
1987a) and in “Titanosauridae indet., DGM Series B” from Brazil (five cervical
vertebrae articulated with 10 dorsal vertebrae;
Powell 1986,
1987a). Thus, titanosaurians are considered to have 13 (Powell 1987a) cervical vertebrae. In
“Titanosauridae indet. DGM Series A,” the neural spines are low and lean
posteriorly in the second to sixth cervical vertebrae (Powell 1986). Based on
comparison of the interprezygapophyseal distance, orientation of the neural
spines, development of the posterior centrodiapophyseal and postzygodiapophyseal
laminae and neural arch fossae, length of the diapophyses, and width/height
ratio of centra with “DGM Series A and B,” Apatosaurus (Gilmore 1936),
Camarasaurus (Osborn and Mook 1921;
McIntosh et al. 1996a,
1996b), and
Haplocanthosaurus (Hatcher 1903), the positions of Malawisaurus
cervical vertebrae were estimated.
In sauropod cervical
vertebrae, the interprezygapophyseal distance, development of laminae and fossae
on the lateral surface of the neural arch, and the length of the diapophyses
increase posteriorly (Osborn and Mook 1921;
Gilmore 1936;
Powell 1986;
McIntosh
et al. 1996a,
1996b). In Haplocanthosaurus (Hatcher 1903), the penultimate
cervical vertebra is wider than high and the last cervical vertebra is shorter
than the preceding cervical vertebra. In Camarasaurus (Osborn and Mook
1921) and Apatosaurus (Gilmore 1936), the lengths of distal cervical
vertebrae are less than the lengths of medial cervical vertebrae. Among titanosaurians, articulated cervical vertebrae are known in “Titanosauridae
indet. DGM Series A” from Brazil, (12 cervical vertebrae, except the atlas,
preserved in articulation with three proximal dorsal vertebrae;
Powell 1986,
1987a) and in “Titanosauridae indet., DGM Series B” from Brazil (five cervical
vertebrae articulated with 10 dorsal vertebrae;
Powell 1986,
1987a). Thus, titanosaurians are considered to have 13 (Powell 1987a) cervical vertebrae. In
“Titanosauridae indet. DGM Series A,” the neural spines are low and lean
posteriorly in the second to sixth cervical vertebrae (Powell 1986). Based on
comparison of the interprezygapophyseal distance, orientation of the neural
spines, development of the posterior centrodiapophyseal and postzygodiapophyseal
laminae and neural arch fossae, length of the diapophyses, and width/height
ratio of centra with “DGM Series A and B,” Apatosaurus (Gilmore 1936),
Camarasaurus (Osborn and Mook 1921;
McIntosh et al. 1996a,
1996b), and
Haplocanthosaurus (Hatcher 1903), the positions of Malawisaurus
cervical vertebrae were estimated.
The anterior ball of
each medial and distal cervical vertebra of Malawisaurus is medially
depressed. The planes of the posterior cups are inclined anterodorsally and not
perpendicular to the axes of the vertebrae, suggesting an anteriorly rising neck
as in Camarasaurus (McIntosh et al. 1996a). The ventral surfaces of the centra are concave medial to the parapophyses but convex posteriorly. Undivided
pleurocoels (sensu
McIntosh 1990) are represented by small fossae
directly ventral to the diapophyses as opposed to multiple large
anteroposteriorly aligned fossae on the centra of other sauropods (Amargasaurus
[MACN-15,
Salgado and Bonaparte 1991]; Apatosaurus [Gilmore 1936];
Brachiosaurus [Janensch 1929a,
1950]; Camarasaurus [BYU
9047; Osborn and Mook 1921;
McIntosh et al. 1996a]; Dicraeosaurus [Janensch 1929b]).
The parapophyses are directed slightly ventrolaterally. Cranial and caudal
peduncle fossae are small. Diapophyses project laterally and become
progressively longer posteriorly.
The neural arches are
low, long, and medially attached anteroposteriorly on the centra. The laminae on
the lateral surfaces of the neural arches are rudimentary (sensu
Wilson
and Sereno 1998). There are only two laminae on the posterolateral surface of a
neural arch, termed the posterior centrodiapophyseal and postzygodiapophyseal
laminae by Wilson (1999). These laminae enclose the infrapostzygapophyseal
fossa. The infrapostzygapophyseal fossae enlarge and deepen posteriorly along
the cervical column. The supradiapophyseal fossae (posterolateral to the
spinoprezygapophyseal laminae and dorsal to the diapophyses) are shallow
anteriorly but deepen and widen posteriorly along the cervical column.
The prezygapophyses
are small and ellipsoidal in anteriorly cervical vertebrae, but are large and
rounded in middle and posterior cervical vertebrae. They extend beyond the
anterior end of the centrum in all cervical vertebrae. The prezygapophyses
become more divergent posteriorly. Intraprezygapophyseal and
intrapostzygapophseal laminae are present in the middle and posterior cervical
vertebrae. The spinoprezygapophyseal laminae are sharp and thin in anterior
cervical vertebrae but thicken posteriorly. Medial to each spinoprezygapophyseal
lamina, at the base, is an ovoid depression in the anterior cervical vertebrae.
The neural spines are
single in all cervical vertebrae. They are rounded and cant posteriorly in the
anterior cervical vertebrae but become progressively more anteroposteriorly
compressed and more vertical posteriorly. In anterior cervical vertebrae, the
neural spines are medial to and are restricted towards the
spinopostzygapophyseal laminae. In the posterior cervical vertebrae, the neural
spines are supported by and rise directly from the spinoprezygapophyseal and the
spinopostzygapophyseal laminae. Mal-245 has well-developed pre-spinal laminae.
Short pre-spinal and long, deep post-spinal fossae occur medial to the
spinozygapophyseal laminae in all cervical vertebrae.
Cervical Ribs.
The cervical ribs (Figure 9) are coossified to the centra (Mal-243, Mal-180,
Mal-245, Mal-278, Mal-280, and Mal-301) or are isolated fragments (Mal-64,
Mal-146, Mal-147, Mal-149, Mal-162, and Mal-187-2). The ribs are nearly parallel
to the long axes of the centra. The heads terminate at the anterior limits of
their associated centra in proximal cervical vertebrae (Mal-180), but terminate
posterior to the anterior ends of their associated centra in medial and distal
cervical vertebrae. This is in contrast to “Titanosauridae indet. DGM Series A”
where the heads of the ribs extend beyond the anterior ends of their associated
centra (Powell 1986,
1987a). The ribs are thin and dorsoventally compressed,
relatively broad transversely in their proximal halves, but thin and rounded
rods in their distal halves. The shafts of the ribs in the proximal and medial
cervical vertebrae extend up to 320 mm beyond the posterior ends of their
associated centra, and even extend beyond the ends of the next succeeding
centra. In the distal cervical vertebrae, the shafts of the ribs do not extend
beyond the centra (Mal-245).
D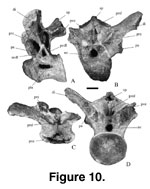 orsal Vertebrae.
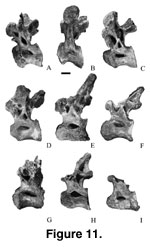
Ten isolated dorsal vertebrae (Figure 9,
Figure 10,
Figure 11,
Table 3) are attributed to Malawisaurus. There are no
hyposphene-hypantrum articular surfaces in any of the vertebrae. The posterior
ends of the centra are flared. The neural arches are low and possess fossae that
vary in size and shape on either side of each vertebra. Undivided and short
neural spines are supported anteriorly by the spinodiapophyseal laminae, not by
the spinoprezygapophyseal laminae as in Apatosaurus (Gilmore 1936), and posteriorly by the spinopostzygapophyseal laminae. All, except Mal-242 (where it
cannot be observed), have well-developed pre-spinal and post-spinal laminae. The
pre-spinal laminae are bifid at the base and are more prominent than the
post-spinal laminae.
orsal Vertebrae.
Ten isolated dorsal vertebrae (Figure 9,
Figure 10,
Figure 11,
Table 3) are attributed to Malawisaurus. There are no
hyposphene-hypantrum articular surfaces in any of the vertebrae. The posterior
ends of the centra are flared. The neural arches are low and possess fossae that
vary in size and shape on either side of each vertebra. Undivided and short
neural spines are supported anteriorly by the spinodiapophyseal laminae, not by
the spinoprezygapophyseal laminae as in Apatosaurus (Gilmore 1936), and posteriorly by the spinopostzygapophyseal laminae. All, except Mal-242 (where it
cannot be observed), have well-developed pre-spinal and post-spinal laminae. The
pre-spinal laminae are bifid at the base and are more prominent than the
post-spinal laminae.
 Among titanosaurians,
the most complete articulated dorsal series known are in a specimen identified
as “Titanosauridae indet. DGM Series B” from Brazil (10 dorsal vertebrae;
Powell
1986, 1987a) and in Opisthocoelicaudia (10 or 11 dorsal vertebrae;
Borsuk-Bialynicka 1977). The description and discussion of
Powell (1986) imply
that “DGM Series B,” Epachthosaurus (MACN-CH 1317, cast), and
Neuquensaurus (Titanosaurus) australis have 10 dorsal
vertebrae, whereas the illustrations of Neuquensaurus (von Huene 1929,
figure 10; Powell 1986, plate 56) suggest the presence of 11 dorsal vertebrae.
Wilson and Sereno (1998, figure 47) indicate that although the number of dorsal
vertebrae in titanosaurians varies between 10 and 12, 10 is the standard number.
Thus, Malawisaurus is predicted to have 10 dorsal vertebrae.
Among titanosaurians,
the most complete articulated dorsal series known are in a specimen identified
as “Titanosauridae indet. DGM Series B” from Brazil (10 dorsal vertebrae;
Powell
1986, 1987a) and in Opisthocoelicaudia (10 or 11 dorsal vertebrae;
Borsuk-Bialynicka 1977). The description and discussion of
Powell (1986) imply
that “DGM Series B,” Epachthosaurus (MACN-CH 1317, cast), and
Neuquensaurus (Titanosaurus) australis have 10 dorsal
vertebrae, whereas the illustrations of Neuquensaurus (von Huene 1929,
figure 10; Powell 1986, plate 56) suggest the presence of 11 dorsal vertebrae.
Wilson and Sereno (1998, figure 47) indicate that although the number of dorsal
vertebrae in titanosaurians varies between 10 and 12, 10 is the standard number.
Thus, Malawisaurus is predicted to have 10 dorsal vertebrae.
In “DGM Series B,” as
well as other sauropods (Hatcher 1903;
Osborn and Mook 1921;
Powell 1986,
1987a;
McIntosh and Williams 1988), the parapophyses move in position from being on the
centrum in proximal dorsal vertebrae to being high on the neural arch in
posterior dorsals. The transverse processes project laterally in the proximal
dorsal vertebrae but dorsolaterally in the distal vertebrae. The
interprezygapophyseal distance progressively decreases posteriorly along the
dorsal series. Comparison with “DGM Series B” and other sauropods (Hatcher 1903;
Riggs 1903;
Osborn and Mook 1921;
Gilmore 1936;
Powell 1986,
1987a;
McIntosh et
al. 1996a;
Jain and Bandyopadhyay 1997) using the positions of the parapophyses,
the orientation of transverse processes, and the interprezygapophyseal
distances, indicates that Mal-181 (Jacobs et al. 1996, figure 5b, c), Mal-236,
Mal-238, Mal-239, and Mal-283 occur in the proximal half of the dorsal series in
positions one to five, while Mal-182 (Jacobs et al. 1996, figure 5d), Mal-237,
Mal-240, Mal-241, and Mal-242 occur in the distal half of the dorsal series, in
positions six to 10.
All the dorsal
vertebrae possess small, anteroposteriorly elongate, eye-shaped (sensu
Calvo and Salgado 1995) pleurocoels that are restricted to the dorsal half of
the centrum. The pleurocoels face laterally in dorsal vertebrae one to three and
dorsolaterally in the other dorsal vertebrae. In Mal-239 and Mal-283, the
parapophyses are subrounded and situated on the centrum. Their ventral limit is
20 mm below the ventral limit of the pleurocoels. Mal-283 is interpreted as the
first dorsal of Malawisaurus, because its parapophyses are immediately
anterior to the pleurocoels, and the diapophyses face ventrolaterally as in the
first dorsal vertebra of other sauropods (Hatcher 1903;
Osborn and Mook 1921;
Powell 1986,
1987a;
McIntosh and Williams 1988). The centrum is shorter and the
zygapophyses are smaller than in the posterior cervical vertebrae but are
similar to the first dorsal vertebra of “Titanosauridae indet. DGM Series A and
B” and of Neuquensaurus (Powell 1986,
1987a).
Parapophyses are
anterior to the pleurocoels in Mal-239. Although the parapophyses are on the
centrum in the second dorsal vertebrae of “Titanosauridae indet. DGM Series A”
from Brazil, the centrum is much shorter than in the first dorsal vertebra
(Powell 1986). The centrum of Mal-239 is much shorter than that of Mal-283
(Table 3) and might be the second dorsal vertebra of Malawisaurus. In
Mal-236, the parapophyses are crescentic and extend from the centrum to the
lower portion of the neural arch. In “DGM Series B,” the parapophyses on the
third dorsal vertebra are on the neural arch and the centrum is wider than high.
The centrum of Mal-236 is similarly wider than high. Mal-236 is also similar to
third dorsal vertebrae of Apatosaurus (Riggs 1903,
Gilmore 1936) and
Camarasaurus (Osborn and Mook 1921) in that half of its parapophysis arises
from the centrum. Thus, Mal-236 might be the third dorsal vertebra of
Malawisaurus.
In all other dorsal
vertebrae, the parapophyses are high on the neural arch. In Mal-238, the
parapophyses are crescentic, and their dorsal limits are slightly lower than the
dorsal limits of the prezygapophyses. In Mal-181 (Jacobs et al. 1996), the parapophyses are at the same level as the prezygapophyses. Parapophyses on the
neural arch, but below or at the same level as the prezygapophyses, occur in
dorsal vertebrae three to five of “DGM Series B.” In Mal-237 and Mal-240, the
parapophyses are subtriangular with medial depressions. The dorsal limits of the
parapophyses are 40 mm higher than the prezygapophyses. In Mal-241, Mal-182, and
Mal-242 the neural arches are not complete, and the parapophyses and diapophyses
are not preserved. However, the decrease in interprezygapophyseal distance
suggests a posterior progression from Mal-181, Mal-237, Mal-241, Mal-240, and
Mal-182 in that order. Mal-242 is considered the most posterior and may be the
last dorsal vertebra of Malawisauru,s because its centrum is the least
excavated among the dorsal vertebrae, similar to the last dorsal vertebrae of
Camarasaurus lewisi (BYU 9047;
McIntosh et al. 1996a) and of
Brachiosaurus (Riggs 1904). Thus, based on parapophyseal position,
interprezygophyseal distance, and size of lateral fossae on the centrum, the
anatomical position of Malawisaurus dorsal vertebrae were approximated.
The prezygapophyses
are at approximately the same level as the postzygapophyses in Mal-283 and in
Mal-239, and are lower than the postzygapophyses in dorsal vertebrae posterior
to the second, Mal-239. Neural arches are attached to the anterior half of
centra in Mal-283, Mal-239, Mal-236, Mal-238, and Mal-181, interpreted
anatomical positions one to five, and are centrally attached to centra in
Mal-237, Mal-241, Mal-240, Mal-182, and Mal-242, interpreted anatomical
positions six to 10. In dorsal vertebrae one to three (Mal-283, Mal-239, and
Mal-236), the posterior limits of the neurocentral junction are anterior to the
posterior limits of the pleurocoels. The neural arch attachments extend the
entire lengths of the pleurocoels in dorsal vertebrae four to nine (Mal-238,
Mal-181, Mal-237, Mal-241, Mal-240, and Mal-182), and extend beyond the
posterior limits of the pleurocoels in dorsal 10 (Mal-242).
The centrodiapophyseal
laminae are dorsally broad and ventrally forked into wide anterior and posterior
centrodiapophyseal laminae in all the dorsal vertebrae except in Mal-242. The
fossae between the anterior and posterior centrodiapophyseal laminae are deep in
Mal-283, Mal-239, Mal-236, Mal-238, and Mal-181, dorsal vertebrae one to five,
shallow in Mal-237, Mal-241, Mal-240, and Mal-182, dorsal vertebrae six to nine,
and have completely disappeared in Mal-242, dorsal 10, so that the
centrodiapophyseal laminae in Mal-242 are massive and not forked. The posterior
centrodiapophyseal laminae and the centropostzgapophyseal laminae are separate
in Mal-283, Mal-239, and Mal-236, dorsal vertebrae one to three, but merge in
all dorsal vertebrae posterior to Mal-236.
The transverse
processes are directed laterally in Mal-238, Mal-239, and Mal-236, and are
slightly dorsolaterally inclined in Mal-283, Mal-181, Mal-237, and in Mal-240.
Diapophyses are ellipsoidal, longer dorsoventrally than anteroposteriorly in
Mal-283, Mal-239, Mal-236, and Mal-238, but longer anteroposteriorly than
dorsoventrally in Mal-181, Mal-237, and Mal-240. The diapophyses face
ventrolaterally in Mal-283, Mal-239, Mal-236, and Mal-238, and laterally in
Mal-181, Mal-237, and Mal-240.
 The neural spine is
paddle shaped in anterior view in each of the anterior dorsals and taper
dorsally in the posterior dorsals. The neural spine lacks lateral pendant
processes. It is inclined posteriorly in any anterior dorsal vertebrae and
becomes vertical posteriorly. Narrow longitudinal fossae occur lateral to the
pre-spinal laminae in Mal-181, Mal-237, and Mal-182, and lateral to the
post-spinal laminae in Mal-238. In other dorsal vertebrae, the presence of
longitudinal fossae on the neural spines cannot be determined because the spines
are incomplete.
The neural spine is
paddle shaped in anterior view in each of the anterior dorsals and taper
dorsally in the posterior dorsals. The neural spine lacks lateral pendant
processes. It is inclined posteriorly in any anterior dorsal vertebrae and
becomes vertical posteriorly. Narrow longitudinal fossae occur lateral to the
pre-spinal laminae in Mal-181, Mal-237, and Mal-182, and lateral to the
post-spinal laminae in Mal-238. In other dorsal vertebrae, the presence of
longitudinal fossae on the neural spines cannot be determined because the spines
are incomplete.
Dorsal Ribs.
Twelve dorsal ribs (Figure 12A-B,
Table 4) are attributed to Malawisaurus.
Comparison with Apatosaurus (Riggs 1903), Camarasaurus (BYU
9047; Osborn and Mook 1921;
McIntosh et al., 1996a;
McIntosh et al. 1996b),
Brachiosaurus (Janensch 1950), and Opisthocoelicaudia
(Borsuk-Bialynicka 1977) suggests that Mal-295, Mal-296, Mal-297, Mal-298, and
Mal-308 occur in the proximal half of the series, from the first to the fifth
position, whereas the remainder occur in the distal half of the series,
posterior to the fifth rib. The posterior surfaces of Mal-282-1 and Mal-282-2,
around positions seven and eight, have quadrangular pneumatic cavities 30 mm
from the capitulum-tuberculum split (Gomani et al. 1999, figures 1d, e)
indicating that pneumatization extends beyond the anterior one third of the
dorsal series mentioned for the Mendoza titanosaurian by
Wilson and Sereno
(1998, p. 52, character 97). The shafts are flattened in all the ribs.
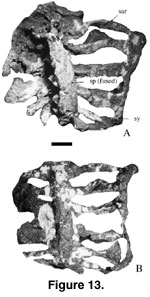 Sacrum. The
sacrum (Figure 13 A-B; anteroposterior length = 574 mm; half width across sacral
two = 310 mm; narrowest half width across sacral three = 266 mm; half width
across sacral six = 395 mm) consists of six vertebrae that are completely fused.
The posterior end of sacral six is not completely preserved. The prezygapophyses
and postzygapophyses are fused. Neural spines are entirely coossified and form a
plate that overhangs the neural arches as in Titanosauridae indet. n. sp. C from
Brazil (Campos and Kellner 1999) and in Epachthosaurus (Powell 1986). The
dorsal surface is roughened with longitudinal grooves. The lateral walls of the
neural spines are thin and strengthened by well-developed laminae as in
Opisthocoelicaudia (Borsuk-Bialynicka 1977). Except for the ribs of the dorsosacral, all the ribs are medially thin, and the laminae are fused to the
parapophyses and diapophyses as in Titanosauridae indet. n. sp. B from Brazil
and other sauropods. The sacral ribs of the dorsosacral are higher on the neural
arch while the ribs of the other sacral vertebrae rise from the ventral surfaces
of the associated sacral vertebrae. The ribs of sacral vertebrae two to five
have subcircular fenestrae proximally as in Saltasaurus (PVL 4017;
Powell
1986). Sacral rib two has an expanded distal ventral plate that overhangs part
of sacral rib three. Ribs two and three lean posteriorly, ribs four and five
lean anteriorly in ventral view, and ribs one and six are vertical. The ventral
openings that separate rib three from ribs two and four are large relative to
other openings. Unlike in the Brazilian sacra (Campos and Kellner 1999), the
distal ends of all sacral ribs in Mal-277-1 are completely coalesced to form the sacricostal yoke as in Opisthocoelicaudia (Borsuk-Bialynicka 1977).
Sacrum. The
sacrum (Figure 13 A-B; anteroposterior length = 574 mm; half width across sacral
two = 310 mm; narrowest half width across sacral three = 266 mm; half width
across sacral six = 395 mm) consists of six vertebrae that are completely fused.
The posterior end of sacral six is not completely preserved. The prezygapophyses
and postzygapophyses are fused. Neural spines are entirely coossified and form a
plate that overhangs the neural arches as in Titanosauridae indet. n. sp. C from
Brazil (Campos and Kellner 1999) and in Epachthosaurus (Powell 1986). The
dorsal surface is roughened with longitudinal grooves. The lateral walls of the
neural spines are thin and strengthened by well-developed laminae as in
Opisthocoelicaudia (Borsuk-Bialynicka 1977). Except for the ribs of the dorsosacral, all the ribs are medially thin, and the laminae are fused to the
parapophyses and diapophyses as in Titanosauridae indet. n. sp. B from Brazil
and other sauropods. The sacral ribs of the dorsosacral are higher on the neural
arch while the ribs of the other sacral vertebrae rise from the ventral surfaces
of the associated sacral vertebrae. The ribs of sacral vertebrae two to five
have subcircular fenestrae proximally as in Saltasaurus (PVL 4017;
Powell
1986). Sacral rib two has an expanded distal ventral plate that overhangs part
of sacral rib three. Ribs two and three lean posteriorly, ribs four and five
lean anteriorly in ventral view, and ribs one and six are vertical. The ventral
openings that separate rib three from ribs two and four are large relative to
other openings. Unlike in the Brazilian sacra (Campos and Kellner 1999), the
distal ends of all sacral ribs in Mal-277-1 are completely coalesced to form the sacricostal yoke as in Opisthocoelicaudia (Borsuk-Bialynicka 1977).
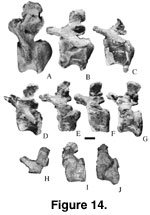 Caudal Vertebrae.
Fifty-one caudal vertebrae (Figure 14,
Figure 15,
Figure 16,
Table 5) of at least three
individuals of Malawisaurus have been described by
Jacobs et al. (1993,
figure 2b, c;
Gomani 1999b, figure 1). The centra are strongly procoelous (sensu
Romer 1956) in the most anterior caudal vertebrae (Mal-191, Mal-225-1,
Mal-232-1, Mal-279-1, and Mal-279-2), becoming slightly procoelous to
platycoelous (sensu Romer 1956) in more the posterior of the anterior
caudal vertebrae (Mal-225-2, Mal-225-3, Mal-225-4, Mal-227, and Mal-228), and platycoelous in the medial and distal caudal vertebrae (sets of Mal-197,
Mal-196, and Mal-192, and isolated vertebrae [Mal-2, Mal-198, Mal-206, Mal-223,
Mal-224, Mal-226, Mal-229, Mal-231, Mal-232-1, and Mal-233]). In the procoelous
tail vertebrae, the posterior ball is restricted to the dorsal half of the
centrum. For a complete description and additional measurements of
Malawisaurus caudal vertebrae see
Gomani (1999b, table 2).
Caudal Vertebrae.
Fifty-one caudal vertebrae (Figure 14,
Figure 15,
Figure 16,
Table 5) of at least three
individuals of Malawisaurus have been described by
Jacobs et al. (1993,
figure 2b, c;
Gomani 1999b, figure 1). The centra are strongly procoelous (sensu
Romer 1956) in the most anterior caudal vertebrae (Mal-191, Mal-225-1,
Mal-232-1, Mal-279-1, and Mal-279-2), becoming slightly procoelous to
platycoelous (sensu Romer 1956) in more the posterior of the anterior
caudal vertebrae (Mal-225-2, Mal-225-3, Mal-225-4, Mal-227, and Mal-228), and platycoelous in the medial and distal caudal vertebrae (sets of Mal-197,
Mal-196, and Mal-192, and isolated vertebrae [Mal-2, Mal-198, Mal-206, Mal-223,
Mal-224, Mal-226, Mal-229, Mal-231, Mal-232-1, and Mal-233]). In the procoelous
tail vertebrae, the posterior ball is restricted to the dorsal half of the
centrum. For a complete description and additional measurements of
Malawisaurus caudal vertebrae see
Gomani (1999b, table 2).
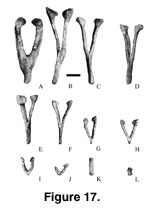
 Chevrons. Among
titanosaurians, only Alamosaurus (25 chevrons;
Gilmore 1946) and
Opisthocoelicaudia (at least 19 chevrons;
Borsuk-Bialynicka 1977) have
complete series of chevrons preserved. Based on comparison with Alamosaurus,
24 recovered chevrons of three different morphologies are considered to belong
to Malawisaurus (Figure 17,
Figure 18,
Table 6). The chevrons belong to at least
two individuals.
Chevrons. Among
titanosaurians, only Alamosaurus (25 chevrons;
Gilmore 1946) and
Opisthocoelicaudia (at least 19 chevrons;
Borsuk-Bialynicka 1977) have
complete series of chevrons preserved. Based on comparison with Alamosaurus,
24 recovered chevrons of three different morphologies are considered to belong
to Malawisaurus (Figure 17,
Figure 18,
Table 6). The chevrons belong to at least
two individuals.
 Mal-192-22, Mal-197-24, Mal-197-25, Mal-197-26, Mal-197-27,
Mal-197-28, and Mal-197-29 were found in natural articulation, and Mal-194,
Mal-197-23, Mal-197-30, and Mal-197-31 were found in close association with an
articulated tail of 21 vertebrae (Mal-197-1 to Mal-197-21;
Gomani 1999a,
1999b) of
Malawisaurus. Mal-195, Mal-277-2, Mal-277-3, and Mal-310 were associated
with the sacrum (Mal-277-1), Mal-255, and the Mal-186 articulated caudal sets.
Malawisaurus is interpreted as having at least 25 chevrons based on
comparison with Alamosaurus (Gilmore 1946). The anteroposterior length of
chevrons decreases to the fifth chevron in Alamosaurus (Gilmore 1946), to
the sixth or seventh chevron in Brachiosaurus (Janensch 1950, figures
109-136), and to the seventh chevron in Opisthocoelicaudia
(Borsuk-Bialynicka 1977).
In these taxa and in Camarasaurus (BYU 9047;
McIntosh et al. 1996a) the length of the haemal canal relative to the length of
the spine increases posteriorly. In Alamosaurus, a V-shaped chevron first
appears between caudal vertebrae 16 and 17 and is referred to as chevron 16
(Gilmore 1946). Thus, in one articulated caudal series (Mal-197) of
Malawisaurus (Gomani 1999a,
1999b), the first V-shaped chevron (Mal-197-26) is
interpreted as the sixteenth chevron.
Mal-192-22, Mal-197-24, Mal-197-25, Mal-197-26, Mal-197-27,
Mal-197-28, and Mal-197-29 were found in natural articulation, and Mal-194,
Mal-197-23, Mal-197-30, and Mal-197-31 were found in close association with an
articulated tail of 21 vertebrae (Mal-197-1 to Mal-197-21;
Gomani 1999a,
1999b) of
Malawisaurus. Mal-195, Mal-277-2, Mal-277-3, and Mal-310 were associated
with the sacrum (Mal-277-1), Mal-255, and the Mal-186 articulated caudal sets.
Malawisaurus is interpreted as having at least 25 chevrons based on
comparison with Alamosaurus (Gilmore 1946). The anteroposterior length of
chevrons decreases to the fifth chevron in Alamosaurus (Gilmore 1946), to
the sixth or seventh chevron in Brachiosaurus (Janensch 1950, figures
109-136), and to the seventh chevron in Opisthocoelicaudia
(Borsuk-Bialynicka 1977).
In these taxa and in Camarasaurus (BYU 9047;
McIntosh et al. 1996a) the length of the haemal canal relative to the length of
the spine increases posteriorly. In Alamosaurus, a V-shaped chevron first
appears between caudal vertebrae 16 and 17 and is referred to as chevron 16
(Gilmore 1946). Thus, in one articulated caudal series (Mal-197) of
Malawisaurus (Gomani 1999a,
1999b), the first V-shaped chevron (Mal-197-26) is
interpreted as the sixteenth chevron.
 Hence, based on comparison with these taxa
and this interpreted position of Mal-197-26, the morphology, proximodistal
length, and the length of the haemal canal relative to the length of the spine,
the anatomical positions of isolated chevrons of Malawisaurus were
estimated. All chevrons lack a bridge of bone so that the haemal canal is open,
unlike the completely enclosed haemal canals or forked chevrons in diplodocids
and dicraeosaurids (Osborn 1899;
Hatcher 1901;
Gillette 1991).
Hence, based on comparison with these taxa
and this interpreted position of Mal-197-26, the morphology, proximodistal
length, and the length of the haemal canal relative to the length of the spine,
the anatomical positions of isolated chevrons of Malawisaurus were
estimated. All chevrons lack a bridge of bone so that the haemal canal is open,
unlike the completely enclosed haemal canals or forked chevrons in diplodocids
and dicraeosaurids (Osborn 1899;
Hatcher 1901;
Gillette 1991).
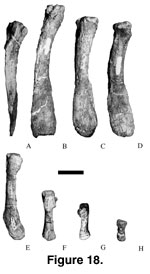 The first morph
(Y-shaped; Figure 17A-F and
Figure 18A-E) has two arms that unite below the haemal
canal to form a spine. All Y-shaped chevrons except Mal-195 have laterally
compressed spines and arms with subequal steep anteroventrally sloping and
gentle anteriorly facing articular facets. In Mal-195 the spine is
dorsoventrally flattened. The heads on the arms curve medially and have a single
articular facet that faces dorsomedially so that when articulated with
vertebrae, it would make an acute angle with the vertebrae column. The spine has
a low, broad median ridge on the posterior surface that divides into two smaller
ridges posteroventrally. The arms are thicker medially than laterally as opposed
to being thicker posteriorly than anteriorly in other Y-shaped chevrons. The
spine is slightly depressed on the anterior surface. Based on these features,
Mal-195 is interpreted as the first chevron. This matches the less developed
chevron articular facet on caudal vertebra Mal-200 as compared to more posterior
vertebrae. In Opisthocoelicaudia (Borsuk-Bialynicka 1977), the spines of
the first four chevrons are dorsoventrally flattened as in Mal-195. A first
chevron that is slightly different from the posterior chevrons also occurs in
Brachiosaurus (Janensch 1950, figure 109) and Mamenchisaurus
hochuanensis (Yang and Zhao 1972).
The first morph
(Y-shaped; Figure 17A-F and
Figure 18A-E) has two arms that unite below the haemal
canal to form a spine. All Y-shaped chevrons except Mal-195 have laterally
compressed spines and arms with subequal steep anteroventrally sloping and
gentle anteriorly facing articular facets. In Mal-195 the spine is
dorsoventrally flattened. The heads on the arms curve medially and have a single
articular facet that faces dorsomedially so that when articulated with
vertebrae, it would make an acute angle with the vertebrae column. The spine has
a low, broad median ridge on the posterior surface that divides into two smaller
ridges posteroventrally. The arms are thicker medially than laterally as opposed
to being thicker posteriorly than anteriorly in other Y-shaped chevrons. The
spine is slightly depressed on the anterior surface. Based on these features,
Mal-195 is interpreted as the first chevron. This matches the less developed
chevron articular facet on caudal vertebra Mal-200 as compared to more posterior
vertebrae. In Opisthocoelicaudia (Borsuk-Bialynicka 1977), the spines of
the first four chevrons are dorsoventrally flattened as in Mal-195. A first
chevron that is slightly different from the posterior chevrons also occurs in
Brachiosaurus (Janensch 1950, figure 109) and Mamenchisaurus
hochuanensis (Yang and Zhao 1972).
The second morph
(V-shaped; Figure 17G-J and
Figure 18F-H) has two arms but lacks the spine. The arms
have small articular facets that face anterodorsally as opposed to the
anteriorly facing articular facets of the Y-shaped chevrons. The third morph
(rod shaped; Figure 17K-L) has two separate arms. The arms are laterally
compressed and have anterodorsal facing articular facets in Mal-152-2 and
Mal-197-31. Mal-197-23 is elongate and cylindrical.
Alamosaurus has
15 Y-shaped chevrons in positions 1 to 15, three V-shaped chevrons in positions
16 to 18, and seven rod-shaped chevrons in positions 19 to 25.
 The articulated
series of Malawisaurus (Mal-197) has five V-shaped chevrons, the last
(Mal-197-30) of which is larger than Mal-152-1. This suggests that Mal-152-1
occurs distal to Mal-197-30 or belongs to a smaller individual. Although the
actual position of Mal-152-1 relative to Mal-197-30 cannot be determined
definitely, Mal-152-1 is considered to be distal to Mal-197-30. It is clear that
Malawisaurus has at least five V-shaped chevrons, more than the three in
Alamosaurus (Gilmore 1946).
The articulated
series of Malawisaurus (Mal-197) has five V-shaped chevrons, the last
(Mal-197-30) of which is larger than Mal-152-1. This suggests that Mal-152-1
occurs distal to Mal-197-30 or belongs to a smaller individual. Although the
actual position of Mal-152-1 relative to Mal-197-30 cannot be determined
definitely, Mal-152-1 is considered to be distal to Mal-197-30. It is clear that
Malawisaurus has at least five V-shaped chevrons, more than the three in
Alamosaurus (Gilmore 1946).
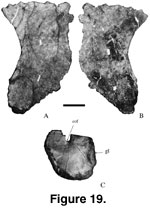
Sternal Plate.
Left (Mal-188-1; Figure 19A-B; length = 490 mm; width = 300 mm; depth range = 5
mm to 50 mm) and right (Mal-188-2) sternal plates were found articulated (Jacobs
et al. 1993, figure 1f). They are semilunar, typical of titanosaurians (Salgado
et al. 1997), and their medial articular surfaces are irregularly ridged. The
distal end of the sternal plate is indented probably for the attachment of the
cartilaginous sternal ribs as in Alamosaurus (Gilmore 1946).
