 Karongasaurus gittelmani sp. nov.
Karongasaurus gittelmani sp. nov.Figure 28
KARONGASAURUS, gen. nov.
Type Species. Karongasaurus gittelmani sp.nov.
Holotype. Mal-175; a dentary with conical replacement teeth; paleontological collection of the Malawi Department of Antiquities.
Etymology. Referring to Karonga District in northern Malawi where the fossil was collected.
Diagnosis. Dentary strongly curved towards symphysis; the rostral end of the dentary is not perpendicular to the long axis of the dentary; dorsal margin of the buccal surface flares outward anteriorly; tooth row restricted to the anterior portion of the jaw; pencil-like teeth with wear facets oblique to the labiolingual axis.
Karongasaurus gittelmani sp. nov.
Figure 28
Type and only species. K. gittelmani sp. nov.
Holotype. A left mandible, Mal-175.
Referred Specimens. Isolated teeth (Table 1; Mal-7, Mal-10, Mal-11, Mal-12, Mal-13, Mal-14, Mal-15, Mal-16, Mal-33, Mal-36, Mal-39, Mal-153, Mal-156, Mal-267, Mal-268, Mal-269, Mal-271, Mal-272, Mal-307).
Diagnosis. As for genus.
Geologic Age and Distribution. Early Cretaceous, Mwakasyunguti area, Karonga District, Malawi, Africa.
Etymology. In honor of Steve Gittelman, friend of science, for his work as president of the Dinosaur Society.
Description
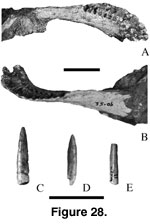
Dentary. The left dentary (Mal-175; Figure 28A–B) is shallow and has 12 tooth positions, even though the symphyseal region is incomplete. The mandible is 138 mm long from the symphysis to the surangular notch and has an 80 mm tooth row. Three unerupted pencil-like teeth are in place. The most posterior tooth position is 62 mm anterior to the surangular notch, almost one third of the distance seen in the mandible of Malawisaurus (Figure 28B; Jacobs et al. 1993). The dorsal margin of the dentary posterior to the tooth row forms a sharp ridge. Anteriorly the dentary is strongly curved towards the symphysis and the dorsal margin of the buccal surface flares outward. The anterior limit of the splenial groove lies below the fourth tooth position from the rear.
Teeth. Slender conical isolated teeth (Figure 28C-E; anteroposterior width range = 30 to 50 mm; average anteroposterior width = 0.4; labiolingual width range = 30 to 40 mm; average labiolingual width = 35 mm) similar to those in the dentary of Karongasaurus are therefore attributed to Karongasaurus. These are more cylindrical than those of Malawisaurus. The crowns have faint anterior and posterior ridges and are more acute cones toward the apex than in Malawisaurus. Wear facets are high angled on the lingual (Mal-153, Mal-272), anterior, and posterior surfaces (Mal-13, Mal-156).
Discussion
The short tooth row and presence of conical teeth in Karongasaurus set it apart from Brachiosaurus, Camarasaurus, Euhelopus, Malawisaurus, Mamenchisaurus, Omeisaurus, and Shunosaurus, all of which have long tooth rows and broad teeth. Karongasaurus shares these characters with Antarctosaurus wichmannianus (von Huene 1929, Powell 1986), dicraeosaurids (Janensch 1935), diplodocids (McIntosh and Berman 1975; Berman and McIntosh 1978), Nemegtosaurus (Nowinski 1971), Quaesitosaurus (Kurzanov and Bannikov 1983), and Rapetosaurus (Curry Rogers and Forster 2001). Unlike dicraeosaurids and diplodocids, where the wear facets on teeth are nearly horizontal, the wear facets on Karongasaurus teeth are steep angled as in Antarctosaurus wichmannianus, Nemegtosaurus, and Quaesitosaurus (Calvo 1994). These taxa have been considered to be either diplodocids or titanosaurians (vvon Huene 1929; Powell 1986; McIntosh 1990; Jacobs et al. 1993; Chatterjee and Rudra 1996; Salgado and Calvo 1997; Sereno et al. 1999; Upchurch 1999). However, Nemegtosaurus, and Quaesitosaurus are considered titanosaurians in the recent analysis of Curry Rogers and Forster (2001) (see also Wilson 2002).
The dentary of Karongasaurus is distinct because the rostral mandibular curvature toward the symphysis is less strong than in Nemegtosaurus, Quaesitosaurus, Antarctosaurus wichmannianus, and diplodocids, in which the mandible is U-shaped in dorsal view. The dentary of Karongasaurus flares outward anteriorly from the inflection point, a feature not observed in other sauropods. The dentary of Rapetosaurus is not known, and therefore cannot be used for comparison.
Postcranial elements found in the Dinosaur Beds lack certain characters (e.g., forked chevrons, deeply cleft neural spines of distal cervical and proximal dorsal vertebrae) that would, if present, suggest diplodocoid affinity (sensu Wilson and Sereno 1998). Therefore, by default stemming from the presence of Malawisaurus, a known titanosaurian, but no other higher level taxon demonstrably present, until more data are found to suggest otherwise, Karongasaurus is considered titanosaurian. If correct, this implies that the possession of slender teeth, the anteromedial mandibular curvature, and the anterior restriction of teeth were characters acquired independently from their presence in diplodocoids.
TITANOSAURIDAE, gen. et sp. indet.
Geologic Age and Distribution. Early Cretaceous, Mwakasyunguti area, Karonga District, Malawi, Africa.
Description
 One small sauropod caudal vertebra
(Mal-5; Figure 29; undesignated morph 1 of
Gomani 1999a, figure 2c) cannot be
assigned to a named taxon with confidence. The centrum of Mal-5 (posterior
fragment) is procoelous (sensu
Romer 1956). A posterior ball is centrally
positioned on the centrum. The ventral surface of the centrum is flat. The
chevron articular facets are poorly developed posteroventral protuberances on
the centrum. The neural arch is not preserved. The width and height of the
centrum, measured from the posterior end, are 38 mm and 53 mm, respectively.
One small sauropod caudal vertebra
(Mal-5; Figure 29; undesignated morph 1 of
Gomani 1999a, figure 2c) cannot be
assigned to a named taxon with confidence. The centrum of Mal-5 (posterior
fragment) is procoelous (sensu
Romer 1956). A posterior ball is centrally
positioned on the centrum. The ventral surface of the centrum is flat. The
chevron articular facets are poorly developed posteroventral protuberances on
the centrum. The neural arch is not preserved. The width and height of the
centrum, measured from the posterior end, are 38 mm and 53 mm, respectively.
The size and the placement of the posterior ball on the centrum of Mal-5, in comparison with strongly procoelous caudal vertebrae of other titanosaurians (SAM 8992; Janensch 1929a; Gilmore 1946; Powell 1986; MUCPv 204; Salgado 1996; Salgado and Calvo 1993; Jain and Bandyopadhyay 1997; Kellner and de Azevedo 1999), suggest that Mal-5 is a middle or a posterior caudal vertebra (Gomani 1999a, 1999b).
Among sauropods, strongly procoelous caudal vertebrae are known in Mamenchisaurus (Young 1958; Yang and Zhao 1972) and in Titanosauria. However, the middle and posterior, but not the posteriormost caudals of Mamenchisaurus are platycoelous. The most posterior caudals of Mamenchisaurus are biconvex. Thus, it is more likely that Mal-5 pertains to Titanosauridae, a family of titanosaurians more derived than Malawisaurus. Because it cannot be assigned to Malawisaurus, it is possible, but not demonstrated, that it may pertain to Karongasaurus.
TITANOSAURIA, gen. et sp. indet.
Geologic Age and Distribution. Early Cretaceous, Mwakasyunguti area, Karonga District, Malawi, Africa.
Description
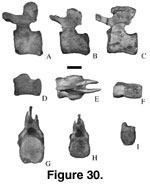 There are five isolated caudal
vertebrae (Figure 30,
Table 9; undesignated morph 2 of
Gomani 1999a, figure 3)
of different morphology from that ascribed to Malawisaurus and to
Titanosauridae indet. (Mal-1 [Jacobs et al. 1993]; Mal-3, Mal-222, and Mal-230
[Gomani 1999a]). The middle (Mal-230) and posterior (Mal-1, Mal-3, Mal-222)
caudal vertebrae of Titanosauria indet. have quadrangular centra that have
nearly flat ventral surfaces, flat lateral surfaces, and poorly developed
chevron articular facets. This contrasts with Malawisaurus in which the
rounded centra have broad, shallow grooves on the ventral surface, slightly
concave lateral surfaces, concave ventral surfaces in lateral view, and
prominent chevron articular facets. It is not clear whether these vertebrae
belong to a different taxon or represent biological or taphonomic variability in
Malawisaurus. If they are determined to be taxonomically distinct, which
appears likely, there are at least three taxa of sauropods present in the
Dinosaur Beds, and it is unclear which of the indeterminate vertebrae, if any,
pertain to Karongasaurus, although some lines of evidence lead to
reasoned speculation (see below).
There are five isolated caudal
vertebrae (Figure 30,
Table 9; undesignated morph 2 of
Gomani 1999a, figure 3)
of different morphology from that ascribed to Malawisaurus and to
Titanosauridae indet. (Mal-1 [Jacobs et al. 1993]; Mal-3, Mal-222, and Mal-230
[Gomani 1999a]). The middle (Mal-230) and posterior (Mal-1, Mal-3, Mal-222)
caudal vertebrae of Titanosauria indet. have quadrangular centra that have
nearly flat ventral surfaces, flat lateral surfaces, and poorly developed
chevron articular facets. This contrasts with Malawisaurus in which the
rounded centra have broad, shallow grooves on the ventral surface, slightly
concave lateral surfaces, concave ventral surfaces in lateral view, and
prominent chevron articular facets. It is not clear whether these vertebrae
belong to a different taxon or represent biological or taphonomic variability in
Malawisaurus. If they are determined to be taxonomically distinct, which
appears likely, there are at least three taxa of sauropods present in the
Dinosaur Beds, and it is unclear which of the indeterminate vertebrae, if any,
pertain to Karongasaurus, although some lines of evidence lead to
reasoned speculation (see below).
![]()