SYSTEMATIC PALEONTOLOGY
Ophidia Brongniart, 1800
Alethinophidia Nopcsa, 1923
Pythoninae Boulenger, 1890
Python
Daudin, 1803
Species indeterminate
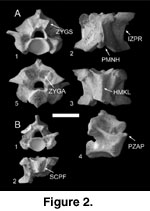
Figure 2
 Referred specimens. H-GSP 13959, 13961-3,
13965, 13968, 14633, 17631, 17720, 19707, 19799, 21005, 21031, 21914, 22078,
22304, 22306, 22309, 23295, 23345, 23723, 24154, 24229, 24350, 24363, 24364,
26723, 27098, 27252, 27719, 27806, 31166, 31318, 31320, 32557, 40204, 40701,
40906, 41161, 41288, 41632, 42267, 46054, 46102, 46184, 46185, 46228, 46420,
46796, 47282, 47381, 47580, 49895-7, 49899, 50044, 50149, 50295, 50425, 50429,
50517, 51109, 51123, 53114, 53115, 53125, 53126, 53178, 53297, 53298, isolated
precloacal vertebrae.
Referred specimens. H-GSP 13959, 13961-3,
13965, 13968, 14633, 17631, 17720, 19707, 19799, 21005, 21031, 21914, 22078,
22304, 22306, 22309, 23295, 23345, 23723, 24154, 24229, 24350, 24363, 24364,
26723, 27098, 27252, 27719, 27806, 31166, 31318, 31320, 32557, 40204, 40701,
40906, 41161, 41288, 41632, 42267, 46054, 46102, 46184, 46185, 46228, 46420,
46796, 47282, 47381, 47580, 49895-7, 49899, 50044, 50149, 50295, 50425, 50429,
50517, 51109, 51123, 53114, 53115, 53125, 53126, 53178, 53297, 53298, isolated
precloacal vertebrae.
Localities and ages.
Y-802 (16.80 Ma), Y-642 (15.20 Ma), Y-478 (14 Ma), Y-650 (13.05 Ma), Y-882 (13
Ma), Y-698 (12.94 Ma), Y-849 (12.81 Ma), Y-750 (12.7), Y-496 (12.30 Ma), Y-634
(12.20 Ma), Y-883 (11.98 Ma), Y-499 (11.95 Ma), Y-515 (11.95 Ma), Y-498 (11.55
Ma), Y-504 (11.52 Ma), Y-061 (11.46 Ma), Y-809 (11.40 Ma), Y-773 (11.34 Ma),
Y-076 (11.31 Ma), Y- 311 (10.00 Ma), Y-262 (9.49 Ma), Y-633 (9.27 Ma), Y-1001
(9.25 Ma), Y-327 (9.18 Ma), Y-401 (8.89 Ma), Y-024 (8.14 Ma), Y-547 (7.93 Ma),
Y-946 (7.76 Ma), Y-457 (7.30 Ma), Y-856 (7.28 Ma), Y-921 (7.24 Ma), Y-370 (7.13
Ma), Y-910 (6.98 Ma), Y-908 (6.78 Ma).
Description. Vertebrae assigned to
Python are large and block-shaped, and include elements from all
recognizable regions of the precloacal vertebrate column. In anterior view
(Figure 2A.1), the cotyle has a strongly subequal ventral margin that is
continuous with the haemal keel, resulting in a slightly pointed cotylar ventral
apex. There is no indication of paracotylar foramina on any recovered specimens
assigned to Python. Dorsal to the cotyle, the neural canal is triangular
and capped by a prominent zygosphene. The zygosphene is tall, with large,
high-angled articular facets. Medially, a pronounced, rounded tuberosity is
present on the anterior face of the zygosphene along the dorsal margin of the
neural canal. Lateral to the cotyle, the prezygapophyses are low-slung and
dorsoventrally thin.
In
dorsal view (Figure 2A.2), the interzygapophyseal ridge possesses a straight
lateral margin that is posteromedially angled between the pre- and
postzygapophyses. The posterior median notch of the neural arch is deep with
straight margins, exposing the posteromedial margins of the zygantral articular
facets in dorsal view. The prezygapophyseal articular facets are ovate, with
straight, transversely oriented anterior margins. Small, pointed accessory
processes are visible at the anterolateral margins of the prezygapophyses.
In
ventral view (Figure 2A.3), the centrum is wide with a thin, poorly defined
haemal keel in all specimens. In vertebrae from the anterior section of the
vertebral column, the hypapophysis originates just anterior to the cotyle.
Hypapophyses are robust and ovoid in cross section. The synapophyses are
moderately developed, with robust, laterally oriented parapophyses and smaller,
posteromedially oriented diapophyses. In vertebrae from the posterior region of
the precloacal column, the paralymphatic channels and subcentral paralymphatic
fossae are well developed and deeply excavated into the ventral surface (Figure
2B.2).
In
lateral view (Figure 2A.4), the interzygapophyseal ridge possesses swollen
lateral margins, and is elevated between the pre- and postzygapophyses. In
posterior view (Figure 2A.5), the lateral margins of the neural arch are
strongly sigmoid in outline. The posterior surface of the arch at the level of
the zygantrum is rugose and pitted for attachment of intervertebral ligaments.
The zygantrum is deep, with a prominent medial ridge.
Discussion. The following character
combination diagnoses referred specimens to Pythoninae: zygapophyseal bridge
straight and posteromedially angled; triangular neural canal; presence of a
zygosphenal tuberosity; absence of paracotylar foramina. Individually, none of
these characters are unique to pythonines (see discussions in
Kluge 1988;
Szyndlar and Rage 2003), but their combined presence is unique to the lineage.
Hoffstetter (1964) recognized Python from the Siwalik Group based on a
single specimen. Generic assignment was based on overall similarity with extant
Python, and Hoffstetter (1964) recognized a specific-level distinction
based on the following characters: relatively elongate centrum (compared to
neural arch width and vertebral height), longitudinal ridge along the haemal
keel, and thick zygosphenal base. There is no indication that a separate
longitudinal ridge can be differentiated from the haemal keel in the Siwalik
specimens, and the morphology of the keel is indistinguishable from that of
other pythonines. A thick zygosphenal base is present in the majority of other
pythonines. The relative length of the centrum varies in the column of all
snakes, including pythonines (Hoffstetter and Gasc 1969;
Polly et al. 2001), and
is not considered diagnostic. Hoffstetter did not name his specimen as a new
species due to limited sample size and the incomplete nature of the specimen.
Based on the generic-level taxonomy of
Kluge (1993b) and large sizes of
specimens, this study also assigns all pythonine specimens to the genus
Python. Because the characters used by
Hoffstetter (1964) to recognize the Siwalik Python as a new taxon are present in extant taxa, or are variable
throughout the vertebral column, I do not make a species-level assignment.
The
Neogene pythonine fossil record includes occurrences in the early Miocene of
Africa, Arabia, Asia, Australoasia, and Europe (Hoffstetter 1964;
Rage 1976,
2003;
Thomas et al. 1981;
Underwood and Stimson 1990;
Ivanov 2000). Geographic
distributions do not exceed the current zoogeography of pythonines (Underwood and Stimson 1990), with the exceptions of a questionable taxon from the Miocene
of Sardinia (Portis 1901), P. euboicus from the early Miocene of
Greece (Szyndlar and Rage 2003), and Python europaeus from the middle
Miocene of France (Ivanov 2000;
Szyndlar and Rage 2003). The earliest
occurrences of Python in the Siwalik section are from localities in the
Kamlial Formation dated at approximately 18 Ma. This record is approximately
coeval with the oldest African and European records, minimally constraining the
timing of pythonine dispersal into mainland Asia and Africa (Kluge 1993b) to no
younger than earliest Neogene.
