 Referred specimens. H-GSP 24402, an
isolated precloacal vertebra.
Referred specimens. H-GSP 24402, an
isolated precloacal vertebra.
Boidae? Gray, 1825
Genus
et species indeterminate
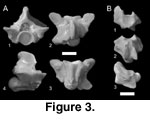
Figure 3A
Referred specimens. H-GSP 24402, an
isolated precloacal vertebra.
Locality and age. Y-039 (approximately 13.5 Ma).
Description. The single precloacal element questionably referred to Boidae indeterminate is incomplete (Figure 3A). The zygosphene, left side of the zygantrum, posterior neural arch, and the distal portions of both synapophyses are not preserved. In all views, the vertebra is short and robust. In anterior view (Figure 3A.1), the prezygapophyses are thin and elongate with low-angled articular surfaces. The prezygapophyseal accessory processes are reduced and do not extend laterally beyond the articular facets. The margins of the cotyle are subequal, with the ventral margin partially eroded. Paracotylar foramina are absent. The preserved portions of the neural canal indicate that it was subtriangular in outline, with the dorsal margin rounded and wide. In dorsal view (Figure 3A.2), the neural spine is anteroposteriorly elongate and transversely thick with a tapered anterior margin. The preserved portions of the posterior median notch indicate that it was well developed. The prezygapophyses extend farther laterally than the postzygapophyses. The interzygapophyseal ridges (Johnson 1955) are strongly concave, with the apex of the concavity anterior to the postzygapophyses. In ventral view (Figure 3A.3), a prominent medial prominence extends along the length of the centrum and is defined by deep paralymphatic fossae (LaDuke 1991). A small haemal keel that includes a small, pointed posterior margin just anterior to the condyle caps the prominence. The paralymphatic fossae extend from the ventrolateral margins of the cotyle anteriorly to the condyle posteriorly. In lateral view (Figure 3A.4), the spinous process is moderately tall and anteriorly sloped. The posterior region of the neural arch is moderately elevated with the lateroventral margin of the neural arch straight and anteroventrally angled with respect to the cotyle.
Discussion. Well-developed paralymphatic fossae indicate that the element is from the posterior region of the precloacal vertebral column (LaDuke 1991). Questionable assignment to Boidae sensu lato is based on the absence of caenophidian characters, including well-developed prezygapophyseal accessory processes and paracotylar foramina. The overall dimensions of the element are additionally similar to those in boines and pythonines. Among fossil taxa, H-GSP 24402 is most similar in general shape to species of the purported boine genus Bavarioboa from the late Paleogene and early Neogene of Europe (Szyndlar and Rage 2003). However, Bavarioboa is not diagnosed by unambiguous apomorphies (Szyndlar and Rage 2003, p. 21), and there are no known postcranial apomorphies that diagnose Boinae sensu stricto. Similarly, cranial characters used to refer Bavarioboa specimens to Boinae (e.g., a posteriorly angled palatine process [Szyndlar 1994, figure 3, V, W]) are either not diagnostic for the clade (Kluge 1991, 1993a, 1993b), or consistently distributed within it (Underwood 1976, figure 4). As a result, there is no definitive fossil record of Boinae in Europe, despite referral of a large number of fossils (e.g., Szyndlar and Rage 2003).
cf.
Erycinae Bonaparte, 1831
Figure 3B
Referred specimens. H-GSP 53417, isolated caudal vertebra.
Locality and age. Y641 (13.55 Ma).
Description. The caudal element is poorly preserved, lacking the majority of the neural arch and pleurapophyses (Figure 3B). Broken edges are worn on the specimen, indicating postmortem hydrologic transport and weathering. In anterior view (Figure 3B.1), the cotyle is circular with equal margins. The lateral margins of the element are partially preserved and are elevated and robust. In ventral view (Figure 3B.2), the bases of the pleurapophyses are preserved on the ventral margin of the lateral body. The pleurapophyses are more laterally offset from the centrum than seen in most taxa. In lateral view (Figure 3B.3), the haemapophyses are triangular in outline with vertical posterior margins anterior to the condyle.
Discussion. Tentative comparison of H-GSP 53417 to Erycinae is based on the presence of highly derived, posterior caudal vertebrae in the clade. Although poorly preserved, the element includes laterally offset pleurapophyses and a robust lateral body without a normally positioned prezygapophysis. These characteristics are not found in the caudal region of the vertebral column in most snakes, but are present in erycines (Sood 1941; Szyndlar 1987). Old World fossil erycines include the genera Albaneryx, Bransateryx, Ogmophis, and Calamagras (Rage 1984; Szyndlar and Rage 2003). The overall shape of H-GSP 53417 is more reminiscent of the comparatively simple caudal elements of Calamagras gallicus (Rage 1977) and Albaneryx (Hoffstetter and Rage 1972) than of the more complex vertebrae of Bransateryx (e.g., Szyndlar 1987).
Booidea (sensu lato) indeterminate
Referred specimens. H-GSP 24381, 24412, 27095, 27103, 27206, 27307, 46027, 46075, 53330, 53334, 53349, 53363, 53386, 53393, 53411, isolated, incomplete precloacal vertebrae.
Localities and ages. Y-747 (18 Ma), Y-641 (13.55 Ma), Y-668 (13.3 Ma), Y-690 (13.04 Ma), Y-825 (12.55 Ma), Y-504 (11.52 Ma), Y-076 (11.31 Ma), Y-797 (11.09 Ma), Y-450 (10.16 Ma), Y-410 (9.29 Ma), Y-024 (8.14 Ma), Y-906 (7.80 Ma), Y-931 (7.24 Ma).
Description. Specimens assigned to Booidea sensu lato (boines, erycines, pythonines) indeterminate consist of fragmentary vertebral centra that lack neural processes. All included elements are small, less than 0.5 cm in length. In ventral view, centra are stout and lack significant ventrolateral cotylar processes in specimens where the cotylar margin is preserved. Ventromedian processes on all specimens consist of poorly defined haemal keels that lack distinct lateral margins and are transversely wider than anteroposteriorly long.
Discussion. Assignment to Booidea is based on the presence of a poorly defined haemal keel, robust vertebral centrum, and absence of ventrolateral cotylar processes. The incomplete nature of specimens considered Booidea indeterminate prohibit refined diagnosis, but they may represent a more extensive record of erycines than can be recognized in this study, as well as neonatal to subadult Python specimens. It is also possible that additional boid or boine taxa are present in the Siwalik sample; however, evaluation of the Old World fossil record of boines does not support such an assignment, as noted above.
![]()