 Referred specimens.
H-GSP 21021, 51107, 50147, incomplete precloacal vertebrae.
Referred specimens.
H-GSP 21021, 51107, 50147, incomplete precloacal vertebrae.
Elapidae Boié, 1827
Bungarus Daudin, 1803
Species indeterminate
Referred specimens. H-GSP 05071, 24365, 24374, 24391; 24396; 27078, 27224, 53026, incomplete precloacal vertebrae.
Localities and ages. Y-450 (10.16 Ma), Y-310 (9.26 Ma), Y-182 (9.16 Ma), Y-388 (8.68 Ma), Y-024 (8.14 Ma), Y-898 (7.77 Ma).
Description and Discussion. All listed specimens are referred to Bungarus on the basis of possessing the elongate, expanded prezygapophyseal accessory processes that are diagnostic for the genus (Hoffstetter and Gasc 1969). Within the genus, all Siwalik specimens are allocated to the polytomy of species more derived than B. flaviceps and B. bungaroides (McDowell 1970; Slowinski 1994).
The first occurrence of Bungarus in the Siwalik Group occurs at approximately 10.2 Ma, and the taxon persists upsection. Implications of Siwalik Group Bungarus for estimating elapid divergence timing and biogeography will be examined elsewhere (Head, in prep).
Elapidae? indeterminate
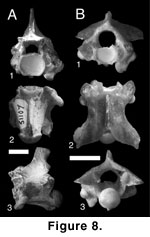
Figure 8A
Referred specimens.
H-GSP 21021, 51107, 50147, incomplete precloacal vertebrae.
Localities and ages. Y-311, (10.00 Ma), Y-908 (6.78 Ma), Y-910 (6.98 Ma).
Description. Three vertebrae represent the largest colubroid specimens recovered from the Siwalik Group. All specimens are incomplete, lacking zygapophyses, the majority of the neural spine, and the majority of the zygantrum and posterior neural arch. In anterior view (Figure 8A.1), the dorsal margin of the zygosphene is flat. The cotyle is circular, and small paracotylar foramina are deeply set along its lateral margins. Small ventrolateral processes are present on the ventral margin of the cotyle, giving it a flattened appearance. Relatively small paralymphatic channels are present between the processes and the synapophyses. The synapophyses possess a low-angled articular surface for the ribs. The diapophyseal articular facet is pronounced and laterally differentiated from the synapophyseal body by a shallow trough. The parapophysis is well developed and rounded.
In ventral view (Figure 8A.2), the centrum is elongate and wide with slightly convex lateral margins. The ventral margin of the cotyle is medially embayed. The hypapophysis is prominent and extends from the cotylar lip to the base of the condyle. The synapophyses are anterolaterally angled away from the centrum. The preserved margins of the interzygapophyseal ridge indicate that it is smoothly convex. In lateral view (Figure 8A.3), the posterior margin of the neural spine is short and much more elevated than the inferred anterior margin. The dorsal margin of the spine appears gently rounded. The ventral margin of the centrum is slightly concave, and the hypapophyseal ventral margin is strongly so. The angle of the hypapophyseal margin indicates that the complete structure was angled posteriorly more than ventrally.
Discussion. Tentative inclusion in Elapidae is based on the large size of the Siwalik specimens, relatively small paralymphatic channels, and concave ventral margin of the centrum. These characters compare well with the king cobra Ophiophagus. A convex ventral margin of the centrum was considered diagnostic of Elapidae by Hoffstetter (1939); however, this character is difficult to assess, and comparison of elapids and other colubroids indicates considerable variability in the curvature of the ventral margin.
Although the referred specimens can be united on the basis of the characters used to compare them with Ophiophagus, they cannot be unambiguously diagnosed to the genus or even to Elapidae. If the specimens do belong to Ophiophagus, then they represent the only fossil occurrence of the genus and a northern geographic range extension. Extant Ophiophagus is not reported from Pakistan (Minton 1966).
Colubridae Opell, 1811
Natricinae
Bonaparte 1838
Genus
et species indeterminate
Figure 8B
Referred specimens. H-GSP 24351, 27164, 50293, incomplete precloacal vertebrae.
Locality and age. Y-547 (7.93 Ma); Y-457, (7.30 Ma).
Description. In anterior view (Figure 8B.1), the vertebrae are low and wide. Paracotylar foramina are present. Elongate ventrolateral cotylar processes form enlarged, circular lymphatic channels between the cotyle and the synapophyses. The parapophyses include large, elongate accessory processes that are rounded in anterior view. The diapophysis is moderately developed relative to other Siwalik taxa. In dorsal view (Figure 8B.2), the neural spine is elongate, uniformly thick, and extends the length of the neural arch. Prominent epizygapophyseal spines are present at the posterolateral margins of the postzygapophyses. The posterior median notch is well developed, with broadly concave margins. In posterior view (Figure 8B.3), the dorsal margin of the neural arch is straight and shallowly angled relative to other taxa. Large parazygantral foramina are present lateral to the zygantral margins. Additional large foramina are present just below the epizygapophyseal spines.
Discussion. Assignment to Natricinae is based on the combination of epizygapophyseal spines and parazygantral foramina. The incomplete nature of Siwalik natricine specimens precludes more refined assignment. There is an extensive list of Old World fossil taxa referred to Natricinae, including Paleonatrix lehmani, P. silesiaca, Neonatrix europaea, N. nova, Natrix longivertebrata, and N. sansaniensis from the Miocene of Europe and Mionatrix diatomeus from the Miocene of China (Sun 1961; Mlynarski et al. 1982; Rage 1984; Szyndlar 1984; Rage and Szyndlar 1986; Szyndlar and Schleich 1993; Ivanov 2000); however, only Paleonatrix is generically diagnosable from vertebral remains by the presence of an apomorphic anteriorly expanded hypapophysis with a square anteroventral margin (Ivanov 2000). No discrete characters have been used in other generic or specific assignments, and there is no basis to allocate Siwalik specimens to previously erected taxa.
![]()