 Hyaenodon sp.:
Qiu et al. 1997, p. 186;
Qiu
et al. 2001, p. 378.
Hyaenodon sp.:
Qiu et al. 1997, p. 186;
Qiu
et al. 2001, p. 378. Order
CREODONTA Cope, 1875
Family
HYAENODONTIDAE Leidy, 1869
Genus
HYAENODON Laizer and Parieu, 1838
Hyaenodon weilini new species
Hyaenodon sp.:
Qiu et al. 1997, p. 186;
Qiu
et al. 2001, p. 378.
Holotype: IVPP V12186, isolated left P4 and M2, right P3, P4, and M2, and root of left C (Figure 2).
Type Locality: IVPP V12186 was acquired from a villager in Zhangjiaping, by Qiu Zhanxiang, Yan Defa, and Xie Junyi on 19 April, 1987, about 10 km north of Lanzhou along the highway to the Lanzhou Airport (Figure 1). Loc. #2 of Zhangjiaping, probably from one of several sand quarries within the first white sandstones in the Middle Member of the Xianshuihe Formation, Zhangjiaping l.f., correlated to magnetic Chron C6n (about 19–20 Ma) in early Miocene (see Qiu et al. 1997; Qiu et al. 2001).
Although the holotype of Hyaenodon weilini was acquired from a farmer living near a “white sands” quarry (see below), our own independent collecting has yielded records (referred specimens) of this species in the “white sands,” increasing our confidence that the holotype is from the “white sands” as well.
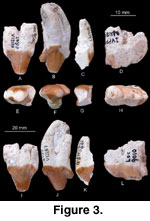 Referred Specimens: IVPP V12187, isolated
left P4 and anterior half of M1, probably from IVPP loc. 8801 in Duitinggou
area, Middle Member of the Xianshuihe Formation; IVPP V12188, isolated left P3,
from IVPP loc. 9512, near the village of Huangyangtou, basal white sandstone in
a sand quarry, Middle Member of the Xianshuihe Formation; and IVPP V12189,
isolated left m1, from IVPP loc. 9010, Yangtonggou, basal white sandstone,
Middle Member of the Xianshuihe Formation (Figure 3).
Referred Specimens: IVPP V12187, isolated
left P4 and anterior half of M1, probably from IVPP loc. 8801 in Duitinggou
area, Middle Member of the Xianshuihe Formation; IVPP V12188, isolated left P3,
from IVPP loc. 9512, near the village of Huangyangtou, basal white sandstone in
a sand quarry, Middle Member of the Xianshuihe Formation; and IVPP V12189,
isolated left m1, from IVPP loc. 9010, Yangtonggou, basal white sandstone,
Middle Member of the Xianshuihe Formation (Figure 3).
Associated Fauna and Age: We here attempt to demonstrate this species to be the last known Hyaenodon in Eurasia, and thus it is a matter of some importance to establish the age of the Middle Member of the Xianshuihe Formation. Vertebrate fossils in the Zhangjiaping local fauna are mostly collected from sediments in and around several light colored coarse sandstones and fine gravels that are concentrated at the lower part of the Middle Member of the Xianshuihe Formation (Qiu et al. 1997; Qiu et al. 2001). Locally known as the “white sands,” in contrast to the “yellow sands” at the bottom of the Lower Member of the Xianshuihe Formation, the channel sandstones are useful as prominent stratigraphic marker beds in geologic mapping, although the sandstones tend to be thinner and less stable laterally toward the upper part of the Middle Member. The Zhangjiaping l.f., first proposed by Qiu (1990) and further elaborated on by Qiu et al. (1997; 2001), includes such early Miocene elements as the primitive ochotonid Sinolagomys pachygnathus and a large-tusked Asiatic rhino Aprotodon lanzhouensis (Qiu and Xie 1997). In particular, the Zhangjiaping l.f. appears to document the first Chinese appearance of proboscideans (see Qiu et al. 2001 for full faunal list). Constrained by the so-called Proboscidean Datum Event (Tassy 1990), Qiu et al. (2001) correlated the “white sands” beds in the Duitinggou section with magnetic Chrons C6n (in part) through C5Cr, approximately 17–20 Ma.
Diagnosis: The Lanzhou creodont is distinguished from the contemporaneous Hyainailouros in having more reduced protocones and metastyle notches in M1-2 with thin shearing blades. Hyaenodon weilini is substantially larger than all known species of Hyaenodon except H. gigas. It differs from the latter in its relatively smaller and lower crowned P4 with more distinct anterior accessory cusp and lingual cingulum, and its distinct anterior ridge on the paracone of M2.
Etymology: In fond memory of Will Downs, whose adopted Chinese name is Dong Weilin, a Chinese approximation of “Downs William,” and who enthusiastically participated in the Lanzhou Basin fieldwork.
Description: Isolated teeth in the holotype IVPP V12186 are regarded as belonging to the same individual because of their consistency of coloration and approximately same stage of wear. However, the possibility exists that they belong to more than one individual because there are moderate size differences between the left and right sides (Table 1).
The P3 is double rooted, and its posterior root is substantially larger than the anterior one, supporting a lingual swelling of the main cusp. The main cusp is slightly recumbent and followed behind by a low posterior accessory cusp. There are faint ridges on the anterior and posterior faces of the main cusp.
The P4 is about the same size as the P3 (in anteroposterior length) and is triple rooted. The lingual root forms the base of a swelled lingual cingulum. This lingual swelling is only slightly more pronounced than that on the P3, and there is no discrete protocone despite the additional root. The P4 main cusp and posterior accessory cusp are more recumbent than in the P3. In addition, the P4 has an incipient anterior accessory cusp that is not present on the P3 (more pronounced in IVPP V12187).
The two upper molars of the holotype are here regarded as M2s because of their considerably larger (anteroposterior length) size than the P4. A prominent M2 is a characteristic of the subfamily Hyaenodontinae, and the M1 is usually about the same size or only slightly larger than the P4. The M2 is highly hypercarnivorous with its long (anteroposteriorly) and thin (transversely) shearing blade and reduced protocone. Although there are three shallow grooves on the labial surface of the M2 (more distinctly displayed on the left M2), none seems to represent a notch between the paracone and metacone, because these grooves extend all the way to the base of the crown. It is likely that the paracone and metacone are fully fused into a single cusp. Instead, the grooves appear to be part of the tooth ornamentation since finer wrinkles are interspersed between the grooves. Wear on the metastyle has eliminated any trace of a metastyle notch (“carnassial notch”). The M2 protocone is extremely reduced (much of its crown was worn off); its existence is only suggested by a small root, which is closely compressed to the larger, labial root below the paracone, and by a shallow notch immediately behind the protocone, which produces a V-shaped enamel edge after wearing. This notch is visible on the left M2. The anterior border of the protocone is behind the anterior ridge of the paracone, and there is a weak indentation between the protocone and paracone. A sharp ridge is present on the anterior surface of the paracone.
A heavily worn m1 (IVPP V12189), measuring 18.5 mm in length and 8.8 mm in width, reveals little more than the presence of a large protoconid-paraconid blade and a small talonid with a single trenchant hypoconid.
Discussion: The Lanzhou Hyaenodon represents the last survivor of this ancient lineage in northern and central Asia. Of the Miocene forms (see Biogeographic below), only Hyainailouros and Hyaenodon achieved the giant size and high degree of hypercarnivory (Helbing 1925; Koenigswald 1947; Ginsburg 1999b) seen in the Lanzhou specimens. Recent phylogenetic analyses suggest that these two genera may represent terminal members of two distinct clades within the family Hyaenodontidae (Polly 1993, 1996), and many of their hypercarnivorous characters (such as the loss of metaconids on lower molars) are independently acquired within the two clades.
Five nominal species of Hyainailouros were recognized by Ginsburg (1980): H. sulzeri, H. bugtiensis, H. fourtaui, H. nyanzae, and H. napakensis. All whose upper cheek teeth are known (including unpublished materials referable to H. bugtiensis from Chinji Formation of Pakistan, Barry, personal commun., 2004) still primitively retain a large (relative to those in Hyaenodon), lingually positioned protocone on the M1-2, and the shearing blade of the M1-2 is still relatively thick (mediolaterally) with a deep metastyle notch. Such primitive morphology of the M1-2 in Hyainailouros is in sharp contrast to the far more hypercarnivorous “carnassials” of Hyaenodon. In the Lanzhou materials, the degree of hypercarnivory far exceeds that seen in Hyainailouros in terms of elongation of shearing blade, reduction of metastyle notch (“carnassial notch”), and reduction of protocone on the M2. The Lanzhou form is thus more readily referable to Hyaenodon.
Numerous species from Africa, Europe, Asia, and North America have been referred to Hyaenodon (37 by Lange-Badré 1979; 42 by Polly 1993). Most of them, however, are from the Eocene to Oligocene and are small to medium-sized predators. By early Miocene time, only a few relics are left. The last North American Hyaenodon (such as H. brevirostris) is found in the early Arikareean (late Oligocene) of California and the northern Great Plains (Mellett 1977; Gunnell 1998; Tedford et al. 2004). None of the North American Hyaenodon achieved the giant size of the Lanzhou taxon. In Europe, the last Hyaenodon are represented by such late Oligocene (Chattian) species as H. milloquensis, H. bavaricus, and H. leptorhynchus. They are small in size, their m1 being in the range of 7-8 mm (see summary in Dehm 1935) in contrast to the 18.5 mm of the Lanzhou Hyaenodon. Three species of Hyaenodon (Isohyaenodon) described by Savage (1965) from the Miocene of Africa, H. andrewsi, H. matthewi, and H. pilgrimi, were based on fragmentary lower jaws (and a single referred M1 of H. pilgrimi). Even the largest of the three species, H. andrewsi (8.9 mm in length), is considerably smaller than the Lanzhou Hyaenodon.
In Asia, Hyaenodon is present in the Paleogene of Mongolia, China, and Kazakhstan (e.g., Matthew and Granger 1924; Teilhard de Chardin 1926; Young 1937; Huang 1982; Dashzeveg 1985; Russell and Zhai 1987; Lange-Badré and Dashzeveg 1989; Huang 1993; Kordikova 1994; Lavrov 1999). Of these, the Mongolian records are the most complete, both in diversity and stratigraphic representation. As summarized by Lange-Badré and Dashzeveg (1989, figure 3), a pattern of declining diversity can be observed from about 3–4 species in the late Eocene to 1–2 species in the Oligocene.
Currently, seven species of Hyaenodon are recognized in the late Paleogene of Asia: H. pervagus Matthew and Granger 1924 from the Loh locality, early Oligocene Hsanda Gol Formation, H. eminus Matthew and Granger 1925 from the late Eocene Ergilin Dzo Formation, H. yuanchensis Young 1937 from middle Eocene Heti Formation of Yuanqu Basin, H. mongoliensis (Dashzeveg 1964) from Khoer Dzan and Bayan Tsav localities of the Ergilin-Dzo Formation, H. incertus Dashzeveg, 1985 and H. (=Neohyaenodon in Lavrov, 1999) gigas Dashzeveg 1985 from Khoer Dzan locality, and H. chunkhtensis Dashzeveg 1985 from Chunkht locality of the Hsanda Gol Formation (see Russell and Zhai 1987 for a summary of these localities).
Only Hyaenodon gigas and H. mongoliensis approach the size of the Lanzhou Hyaenodon. H. mongoliensis is based on lower jaw and teeth only, and thus cannot be directly compared with the Lanzhou materials. Given that H. gigas and H. mongoliensis co-occur in the Khoer Dzan locality of the Ergilin-Dzo Formation, and even allowing that these two giant Hyaenodon species are distinct, it is likely that they are closely related (either sister species or anagenetically successive forms). If so, the following substantial differences of H. gigas from H. weilini also likely to apply to H. mongoliensis.
Based on Dashzeveg’s (1985, figure 12) illustration, the P4 of Hyaenodon weilini differs from H. gigas in the following features: somewhat smaller M2 (Table 1), substantially smaller P4 (i.e., relatively small P4 compared to M2), and P4 with more distinct anterior accessory cusp and a lingual cingulum. Perhaps the most prominent difference between these two species is the considerably higher crowned P4 in H. gigas (see especially Dashzeveg 1985, figure 12b). Considering the fact that PSS 27-10, holotype of H. gigas, has suffered far more wear on the tip of the main cusp of the P4, the crown height of PSS 27-10 may be twice as tall as that of IVPP V12187. The P4 in PSS 27-10 shows substantial apical wear, in contrast to the shearing wear on the M1–2, indicating its primary function as a bone-crushing tooth. The M2 has gone through extensive wear on PSS 27-10 and shows signs of lingually bent shearing blades typical for many late adult individuals of Hyaenodon. PSS 27-10 appears to lack a distinct anterior ridge on the anterior face of the paracone, which is present in IVPP V12186. This ridge is present in earlier and more primitive species from North America (such as in some individuals of H. vetus, H. montanus, and H. horridus, see Mellett 1977, figures 6, 7, 10), and its absence in H. gigas may be another derived character.
A new genus and species, Macropterodon zelenovi Lavrov 1999, was named on a lower jaw fragment, PIN 3110/630, also from the Khoer Dzan locality in the Ergilin Dzo Formation. This taxon is mainly diagnosed on the basis of a large, high-crowned p4 that is relatively larger than the m1. Although lacking comparable materials of Hyaenodon gigas and Macropterodon zelenovi, we cannot help but note the similar giant size and robustness, as well as a similarly massive P4 in both species. The possibility of that these two species are synonymous should be explored when additional materials become available.
The size difference between the P3s in IVPP V12186 (25.6 x 18.0 mm) and V12187 (21.1 x 14.3 mm) is substantial (more than 20%), but is well within the documented variation (16 to23 mm for P3s, or more than 40%) in Hyaenodon horridus, the largest species in North America (Mellett 1977). At the present our sample size is too small to evaluate intraspecific variations and we treat this size difference as variation within species.
Overall, the above outlined features seem to indicate a somewhat more primitive stage of evolution in Hyaenodon weilini, and considering the long hiatus (more than 10 m.y.) between H. weilini, on the one hand, and H. gigas and H. mongoliensis, on the other, it seems unlikely that the Lanzhou Hyaenodon is conspecific with the Ergilin Dzo forms. Instead, it represents the last survivor of this genus in Eurasia, one that developed gigantism independently from its earlier Asian relatives.
Hyaenodon pervagus Matthew and Granger, 1924
Material: IVPP V13573, an isolated left p4, from IVPP loc. 9016, 100 m west of the village of Duitinggou from basal yellow conglomeratic sandstones, Lower Member of the Xianshuihe Formation, Nanpoping local fauna, in magnetic Chron C11r (about 30.3–30 Ma) in late early Oligocene (Qiu et al. 2001).
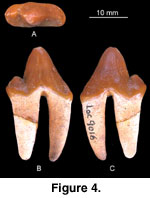 Comments: IVPP V13573 (Figure 4) has a tall,
slightly recumbent main cusp, a thin anterior ridge along its anterior face, and
a posterior ridge reaching down to the posterior accessory cusp. The posterior
accessory cusp is well developed. A very weak lingual cingulum is present on the
anterior and posterior half of the tooth.
Comments: IVPP V13573 (Figure 4) has a tall,
slightly recumbent main cusp, a thin anterior ridge along its anterior face, and
a posterior ridge reaching down to the posterior accessory cusp. The posterior
accessory cusp is well developed. A very weak lingual cingulum is present on the
anterior and posterior half of the tooth.
Measuring 17.4 x 7.2 mm, IVPP V13573 is nearly identical in size to the holotype of Hyaenodon pervagus Matthew and Granger 1924 (17.3 x 8.4 mm) from the Hsanda Gol Formation, although the latter falls in the upper range of the species (Lange-Badré and Dashzeveg 1989, table 2). The size and shape of IVPP V13573 easily distinguish it from other Hsanda Gol species (p4 of H. chunkhtensis measuring 7.0 x 3.0 mm, p4 of H. eminus measuring 11.0 x 5.5 mm, and H. incertus having a robust and broad p4 measuring 17 x 11 mm).
![]()