DISCUSSION
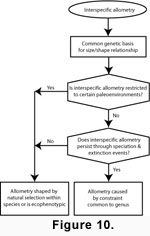 Determining the cause of interspecific allometry in Caryocorbula requires a series of inferences
(Figure 10). The consistent correlation of size and shape in
Caryocorbula that is not seen in its sister clade (Bothrocorbula +
Hexacorbula), implies that this relationship is genetically based and is an emergent property of
Caryocorbula as a whole. Further, because a clear and persistent allometric pattern is present in a data set where paleoenvironmental conditions were neither constant nor sampled consistently over geologic time, neither ecophenotypic plasticity nor natural selection can be the cause of
Caryocorbula allometry. Finally, a role for constraints in this interspecific allometry is indicated because this trend persists through multiple speciation and extinction events (see
Gould 1966;
Maynard Smith et al.
1985).
Determining the cause of interspecific allometry in Caryocorbula requires a series of inferences
(Figure 10). The consistent correlation of size and shape in
Caryocorbula that is not seen in its sister clade (Bothrocorbula +
Hexacorbula), implies that this relationship is genetically based and is an emergent property of
Caryocorbula as a whole. Further, because a clear and persistent allometric pattern is present in a data set where paleoenvironmental conditions were neither constant nor sampled consistently over geologic time, neither ecophenotypic plasticity nor natural selection can be the cause of
Caryocorbula allometry. Finally, a role for constraints in this interspecific allometry is indicated because this trend persists through multiple speciation and extinction events (see
Gould 1966;
Maynard Smith et al.
1985).
As outlined in the introduction, constraint studies lack a common terminology. We have chosen to use the recent classification of
Richardson and Chipman (2003) as a basis for discussion of constraint in
Caryocorbula. Within this classification (Table
1), the most likely mechanisms to explain the interspecific allometry in
Caryocorbula are either historical (a type of generative constraint) or horizontal (a selective constraint) constraints.
Richardson and Chipman (2003)
define historical constraints as generative constraints in which the evolutionary history of an organism limits the developmental pathways that it can evolve, either because of current form (e.g.,
Resnik 1995; but see
Schwenk
1995) or because of the patterning mechanisms available to it (e.g., Gould
1989).
Caryocorbula, and perhaps other corbulids, may be subject to historical constraint through their characteristically inequivalved condition. In
corbulids, the smaller left valve fits partially to completely into the right valve. One way that the inequivalved condition may constrain morphology is by limiting internal space. In fact, we see a pattern consistent with this limitation in our data. Most
Caryocorbula species (and most corbulid species for that matter) have relatively inflated valves. In our dataset, only two
Caryocorbula species have uninflated valves (Caryocorbula sp. B and
Caryocorbula sp. C). Perhaps not coincidentally, these two species are also the ones that do not fit into the overall allometric trend
(Figure 7).
In addition to limits on internal space, historical constraints acting through the ontogeny of valve accretion in corbulids may play a significant role in determining valve shape.
Many corbulids experience an initial period of valve-edge accretion producing a very thin shell
(Goodwin et al.
2003). In this initial phase, left and right valves can be nearly
equivalved, even in highly inequivalved genera such as Varicorbula (Anderson
1996). This valve-edge accretion phase is followed by a phase where accretion occurs predominantly onto valve inner surfaces, making the growth trajectory highly oblique
to perpendicular to the commissure plane
(Wrigley 1946;
Goodwin et al.
2003). This change in growth direction may accompany the development or accentuation of the inequivalved condition. Adult valve shape may be constrained, therefore, by the need for functional valve articulation between valves that differ in size and shape in the latter phase of this complex accretionary history.
It is also possible that horizontal constraints (Richardson and Chipman
2003), in which some developmental pathways influence many different processes in development so that changes in any pathway are likely to have a negative selective value because of pleiotropic effects (see also
Raff
1996). For instance, selection may be acting on
Caryocorbula size, but if shape is linked to size by pleiotropy, an interspecific allometric trend such as we document would result (see
Gould 1977;
Gould and
Lewontin 1979; Cheverud 1984;
Norris
1991). In previous work, we have called on regional changes in nutrient availability as a cause of dramatic size trends in corbulid and chionine bivalves of the Caribbean Neogene (e.g.,
Roopnarine 1996;
Anderson
2001). More specifically, we have inferred that speciation and extinction of large-bodied members of these clades are controlled by changes in nutrient availability and primary productivity. Therefore, for
Caryocorbula, environmental conditions (i.e., increased nutrient availability) could allow the evolution of large-bodied species, but constraints could restrict the species that arise to a narrow range of shapes.
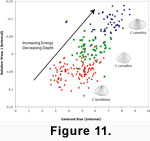 Although interspecific allometry in Caryocorbula may be a byproduct of constraint, this allometric trend, nonetheless, can have adaptive significance
(Gould and
Lewontin 1979; Maynard Smith et al. 1985;
Norris 1991;
Amundson 1994;
Levinton
2001). In other words, constraint may channel morphologic variability but natural selection nevertheless acts as the underlying force for change
(Bell 1987;
Gould 1989;
Landman 1989;
Geary et al.
2002). In fact, evidence that the correlation of size and shape are adaptive in
Caryocorbula can be inferred through examination of areas where precise paleoenvironmental data are available, such as Neogene deposits of the Dominican Republic
(Figure 11).
Although interspecific allometry in Caryocorbula may be a byproduct of constraint, this allometric trend, nonetheless, can have adaptive significance
(Gould and
Lewontin 1979; Maynard Smith et al. 1985;
Norris 1991;
Amundson 1994;
Levinton
2001). In other words, constraint may channel morphologic variability but natural selection nevertheless acts as the underlying force for change
(Bell 1987;
Gould 1989;
Landman 1989;
Geary et al.
2002). In fact, evidence that the correlation of size and shape are adaptive in
Caryocorbula can be inferred through examination of areas where precise paleoenvironmental data are available, such as Neogene deposits of the Dominican Republic
(Figure 11).
In these late Miocene and early Pliocene sediments, valve morphology, sediment type, and inferred water depths are closely related, and this relationship cannot be explained by taphonomic processes such as transport and size sorting
(Anderson 1994,
1996). Valves of the relatively large
C. caimitica occur in marginal- to shallow-marine sands and silts with evidence of sediment movement and where bioclasts are concentrated into lenses, burrows, or lenticular beds. Such strata with concentrated bioclasts were probably deposited in higher energy conditions in which
C. caimitica would be less susceptible to disinterment because of its larger size, and would be capable of more efficient and deeper burial because of its elongate shape and more prominent rostrum
(Stanley
1970). The morphologically intermediate
C. cercadica also is found in shallow-marine deposits, but those formed in a lower energy setting, as indicated by the species' occurrence in lenticular beds rich in bioclasts with a silty matrix. Finally, valves of the small species
C. lavalleana typically occur in deeper-water deposits of finer-grained sediment where bioclasts are dispersed through the matrix, indicating low energy conditions. The triangular shape of
C. lavalleana would increase its buoyancy within these softer sediments.
Further evidence to support an adaptive explanation for valve shape may be gleaned from
Caryocorbula sp. B and Caryocorbula sp. C, which do not fall within the allometric trend
(Figure 7). These species have valve outline shapes typical of species with smaller body sizes. These un-elongated moderate-sized species may have compensated for a lack of anterior-posterior streamlining with a thinner profile provided by their relatively uninflated valves.
