INTRODUCTION
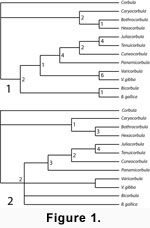 Species of Caryocorbula, one of the most abundant genera of molluscs in the Neogene of the Caribbean
(Jackson et al.
1999), typically have been identified using qualitative descriptions of valve outline shape and valve size. This genus, however, is relatively conservative in its morphology, making qualitative methods for alpha-level taxonomy difficult. Using multivariate analyses of landmark data, we examined the relationship of size and shape within the clade
Caryocorbula + (Hexacorbula + Bothrocorbula) (Figure 1,
Figure 2) and within
Caryocorbula to document patterns of morphologic evolution in these taxa, focusing on the Caribbean
Neogene. We found that Caryocorbula exhibits a strong interspecific allometric trend through the Neogene in this region; a trend that persists through speciation and extinction events, and across environmental gradients. We see roles for both constraint and selection in shaping these morphologic patterns.
Species of Caryocorbula, one of the most abundant genera of molluscs in the Neogene of the Caribbean
(Jackson et al.
1999), typically have been identified using qualitative descriptions of valve outline shape and valve size. This genus, however, is relatively conservative in its morphology, making qualitative methods for alpha-level taxonomy difficult. Using multivariate analyses of landmark data, we examined the relationship of size and shape within the clade
Caryocorbula + (Hexacorbula + Bothrocorbula) (Figure 1,
Figure 2) and within
Caryocorbula to document patterns of morphologic evolution in these taxa, focusing on the Caribbean
Neogene. We found that Caryocorbula exhibits a strong interspecific allometric trend through the Neogene in this region; a trend that persists through speciation and extinction events, and across environmental gradients. We see roles for both constraint and selection in shaping these morphologic patterns.
Allometry and Constraint
Allometry can be defined as differential growth of body parts as the size of an organism increases.
Klingenberg (1998) noted that allometric studies could be divided into two schools with different emphases. Studies in what he terms the
Huxley-Jolicouer School are concerned with patterns of size covariation among several morphologic traits, in particular size covariation that fits the equation of simple allometry
(y=bxa where a is a constant but does  one (1) if the two traits are measured on the same scale). In such studies, shape is of peripheral importance.
one (1) if the two traits are measured on the same scale). In such studies, shape is of peripheral importance.
 Allometric studies in the
Gould-Mosimann School focus on the relationship of shape and size, and give simple allometry no special status (i.e., allometry represents any type of non-isometric growth), except to note that it often fits empirical data well (e.g.,
Gould 1966,
1975,
1977;
Jungers et al.
1995).
Allometric studies in the
Gould-Mosimann School focus on the relationship of shape and size, and give simple allometry no special status (i.e., allometry represents any type of non-isometric growth), except to note that it often fits empirical data well (e.g.,
Gould 1966,
1975,
1977;
Jungers et al.
1995).
Allometry is often categorized by the types of data collected for study (see
Cock 1966;
Gould
1966). With static or intraspecific allometry, the covariation of traits among individuals within a particular ontogenetic stage of a single species is recorded. Ontogenetic allometry involves the determination of covariation among traits across ontogenetic stages of given species (but not necessarily within one individual, which is termed longitudinal allometry). The allometry tracked in this study is interspecific allometry, defined as allometry among species of different sizes within a single clade at the same growth stage (i.e., adult).
Broadly defined, constraints are mechanisms that limit or channel adaptive response in adult body plans, and allometry and morphologic constraints have been linked by a number of authors. For instance,
Maynard Smith et al. (1985) note that allometric relations among adults of different species
(interspecific allometry) that resemble growth stages of a single species (ontogenetic
allometry) are strongly suggestive evidence of constraint. Further, Gould (1966) notes that interspecific allometry (as we document here for
Caryocorbula) is the most likely type of allometry to illustrate change that must occur as size increases (i.e., shape that is functionally constrained by size).
Although discussed widely, constraint terminology has defied attempts at standardization, with categories and definitions of constraints varying among authors. For instance, phyletic constraints have been defined in related but non-interchangeable ways as adaptations retained in new ecologic settings (i.e., phyletic inertia;
Gould and
Lewontin 1979) and as "establishment of a particular ontogeny to the exclusion of other options"
(Raff
1996). Further, a similar concept of historical constraint has been defined as inherited allometries channeling evolutionary change
(Gould
2002), as the form of ancestral species setting the course of evolutionary change
(Thomas and Rief
1993), and as the evolutionary history of an organism limiting the developmental pathways that can evolve
(Richardson and Chipman
2003).
As is apparent from these definitions, constraint terminology also includes a mix of pattern definition and process inference, and is often vague about causal mechanisms, primarily because these mechanisms are difficult to determine, even within living species. In addition, processes underlying specific constraints may be recategorized from one author to the next, so that pleiotropy, for example, has been listed as a mechanism of developmental constraint
(Cheverud
1984), genetic constraint (Gould and
Lewontin 1979, Atchley and Hall 1991,
Schwenk 1995,
Schlichting and Pigliucci
1998), and horizontal constraint (Richardson and Chipman
2003).
The types of mechanisms considered to be constraints are the topic of considerable discussion. For example, the most widely quoted constraint definition is that of
Maynard Smith et al. (1985) who defined developmental constraints as "biases in production of variant phenotypes, or limitations on phenotypic variability caused by the structure, character, composition or dynamics of the developmental system." A number of authors have attempted to narrow this definition, including
Fusco (2001) who, using the term reproductive constraint, defined constraint as a "bias on the production of new ontogenetic trajectories caused by current mechanism of transmission of hereditary factors and laws of epigenetic interaction." In this definition, only phenomena that bias the organization of new ontogenetic trajectories are included, with phenomena that are properly natural selection acting during development excluded (see also
Schlichting and Pigliucci
1998). Alternatively, Richardson and Chipman (2003) draw a distinction between what they call generative and selective constraints but consider both as types of constraint
(Table 1). In this classification, generative constraints limit production of certain phenotypes during ontogeny and result in non-random production or nonproduction of variants. Selective constraints arise by natural selection acting during embryonic development, and remain approximately constant through a broad range of environments.
Clearly, the classification of constraint's causal mechanisms remains in flux. We do not attempt here to make further contributions to this situation. Instead, in the discussion below we will examine the possible role of constraint, and causal mechanisms of constraint, in the morphologic evolution of
Caryocorbula using the broadly inclusive but clearly defined classification developed by
Richardson and Chipman
(2003).
