MATERIALS AND METHODS
Phylogenetic Analyses
Phylogenetic analyses, which are described in detail in
Anderson and Roopnarine
(2003), are only briefly outlined here. We used 24 fossil and Recent species to construct character states of both composite and exemplar taxa for phylogenetic analyses of Neogene genera and subgenera of the Corbulidae from tropical America. All characters used in the analyses were
conchologic, and character polarity was determined using Corbula as an
outgroup. We ran analyses on two matrices: one of 73 and the other of 70 characters, which differed in the manner that
commarginal-rib characters were coded. Phylogenetic analyses using branch and bound searches and maximum parsimony were completed in
PAUP* 4.0b10 (Swofford
2002).
We used Bremer decay indices (Bremer
1994) to compare the robustness of cladogram nodes. This decay index tracks the survival of nodes as the length of accepted cladograms increases incrementally. In other words, one computes a strict consensus tree incorporating the shortest
tree(s) and those one step longer, and notes which nodes retain their resolution. The process is repeated incrementally until all resolution is lost on the strict consensus tree.
Morphometric Analyses
We used landmark-based geometric morphometrics to quantify size and shape variation
(Bookstein
1991). Geometric morphometrics are used increasingly for morphometric studies because the methods allow 1) complete separation of size and shape into distinct variables, 2) segregation of shape into uniform and non-uniform components (see below), and 3) powerful means to visualize morphologic differences using thin plate spline
(tps) techniques. Further, the shape variables derived from geometric morphometric analyses can be incorporated into commonly used multivariate analyses including analysis of covariance
(ANCOVA) (Rosas and Bastir
2002), multivariate regression (Zelditch and Fink 1995;
Penin and Berge 2001;
Rosenberg
2002), principal components analysis (Zelditch et al.
2003), and canonical variates analysis (multivariate version of discriminate analysis; this study). In fact, a number of authors have used geometric morphometrics in allometric studies both within species
(Zelditch et al. 1993;
Zelditch and Fink 1995;
Rosas and Bastir
2002) and among species (Fink and Zelditch 1995;
Penin and Berge 2001;
Rosenberg
2002;
Zelditch et al.
2003).
In geometric morphometrics, ordinary Cartesian coordinates of geometrically homologous points (i.e., landmarks) are used to define the distributions of specimens in Kendall shape space around a Procrustean distance-minimized common reference form
(Rohlf
1996). Specimen deviation from this consensus form is then calculated as the location of the specimen on a series of principal warp vectors (= partial warp scores). The principal warps are the principal axes of a normal Euclidean space tangential to the curved Kendall shape space, and are termed the non-uniform component of shape description because they summarize localized shape variation at variable geometric scales. The projection of the shape space distributions (i.e., partial warp scores) into Euclidean space is a mathematical convenience that permits their use in normal univariate and multivariate analyses. An additional component of shape description is the uniform or affine transformations of global stretching/compressing and shearing. Together, the uniform and non-uniform shape components provide a complete description of specimen shape.
In geometric morphometric studies, centroid size, the square root of the sum of squared distance of a series of measured landmarks to their common
centroid, is often used as an estimate of overall size. Centroid size is a desirable measure of geometric scale because it is the only scaling variable that is uncorrelated with shape measures in the absence of allometry
(Bookstein
1991). It, therefore, presents a clean and interpretable separation between organismal shape and size.
As part of this study, we defined 21 morphospecies of
Caryocorbula represented by 1,145 specimens and used these species, along with 4 species of
Bothrocorbula and Hexacorbula represented by 134 specimens, in a series of geometric morphometric analyses. Samples used were from Neogene deposits of Venezuela, Panama, Costa Rica, Dominican Republic, and Jamaica, as well as samples of Holocene corbulids collected in the Caribbean and Gulf of Mexico
(Table 2, Table 3,
Appendix). Collections acronyms are as follows:
NMB—Naturhistorisches Museum, Basel, Switzerland, and TU—collections of Harold and Emily Vokes (now housed at the Paleontological Research Institution, Ithaca, New York and the Florida Museum of Natural History, Gainesville, Florida).
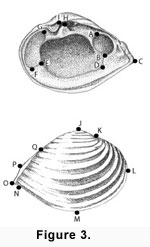 Because corbulids are inequivalved, we used only right valves in analyses, but captured landmarks of both internal and external views of specimens
(Figure 3). Traditional species-level diagnostic characters within these genera are valve outline shape and valve size. Expression of ribs, and the lunular pit in
Bothrocorbula, are used to a lesser extent for species discrimination. External landmarks generally track valve outline shape directly. The internal landmarks used are only indirectly related to traditional species-level features, although the relative placement of internal features is influenced by valve shape. Most internal landmarks, however, can be registered with greater precision, because more represent either the intersection of features, or maxima of curvature on very tight curves
(Figure 3).
Because corbulids are inequivalved, we used only right valves in analyses, but captured landmarks of both internal and external views of specimens
(Figure 3). Traditional species-level diagnostic characters within these genera are valve outline shape and valve size. Expression of ribs, and the lunular pit in
Bothrocorbula, are used to a lesser extent for species discrimination. External landmarks generally track valve outline shape directly. The internal landmarks used are only indirectly related to traditional species-level features, although the relative placement of internal features is influenced by valve shape. Most internal landmarks, however, can be registered with greater precision, because more represent either the intersection of features, or maxima of curvature on very tight curves
(Figure 3).
Centroid size was computed for both internal and external views using
Shape (Cavalcanti
1996). However, only centroid size based on internal landmarks are used to illustrate allometry because the measure is based on more precisely placed landmarks (and an additional landmark) than external centroid size. Both uniform and non-uniform (partial warp) scores were computed using the program
tpsRelw (Rohlf
2002) with scores for internal and external landmarks calculated separately. We computed scores for all three genera and for
Caryocorbula only. For each data set, we examined the homogeneity/heterogeneity of uniform and non-uniform scores using relative warp analysis, a modified principal components analysis. For
Caryocorbula, we used these results as a first step in defining
morphospecies. Group identifications (among genera and among Caryocorbula species) were tested further with canonical variates analysis
(CVA) of the same scores. In preliminary analyses, CVAs in which scores of internal and external landmarks were combined produced better discrimination among identified groups than did CVAs where internal and external landmarks were analyzed separately. Therefore, with one exception, these combined results were used to visualize morphologic variability in our datasets. For
Caryocorbula, initial CVA results were used to test and refine original morphospecies interpretations, which were then used in the CVAs reported here.
