 Referred specimens. PTRM 1829, 2073, 5629, 5633, 5657 (posterior presacral vertebra,
Figure 21), 5663
Referred specimens. PTRM 1829, 2073, 5629, 5633, 5657 (posterior presacral vertebra,
Figure 21), 5663
A great many "lizard" vertebrae are known from V89002. One morphotype deserves special attention. Its description separate from the taxa described above is not intended to imply that it represents a different taxon, only that their attribution to a particular taxon is not yet fully warranted.
Referred specimens. PTRM 1829, 2073, 5629, 5633, 5657 (posterior presacral vertebra,
Figure 21), 5663
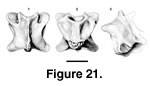
Description. PTRM 5657 is a well-preserved posterior presacral vertebra. The neural arch surrounds a large, rounded neural canal. The neural crest is low anteriorly but rises posteriorly into a moderately developed, posteriorly inclined spine (Figure 21.1,3). The anterior border of the neural arch is strongly concave. Posteriorly, the neural spine is mirrored on the underside of the neural arch by a small ridge that does not extend into the neural canal. Accessory articulations (zygosphenes and zygantra) are well developed. The zygosphenes are oriented ventrolaterally, at an angle of ~40° to the prezygapophyses. The zygophenes are continuous with the prezygapophyses; that is, there is no notch separating zygosphene from prezygapophysis (Figure 21.3). The articular surfaces of the pre- and postzygapophyses are rounded (Figure 21.1–2). There is a pair of tiny foramina below the prezygapophyses on the anteromedial edge of the neural arch.
On the ventral surface are well-developed, bilateral subcentral impressions, giving rise to a mid-sagittal ridge (Figure 21.2). The ridge is not rounded but rather squared off in cross-section, and a narrow, shallow, longitudinal groove runs along its length. In other words, the ridge appears weakly paired (Figure 21.2). No subcentral foramina are present. Ridges of bone run posteroventrally from the small, rounded synapophyses toward the condyle, bounding the subcentral impressions laterally. The centrum is moderately depressed. Half of the specimens referred here have a paired ventral ridge (PTRM 5629, 5657, 5663).
Remarks. It is tempting to try to refer these lizard vertebrae to a species described above (since they were surely produced by one of them). The largest iguanids in the locality are Polychrus charisticus, Iguanids MPH-1 and -2, and cf. Aciprion sp. P. charisticus, Iguanid MPH-1, and cf. Aciprion sp., represented by 12, 9, and 8 specimens (respectively), are also the most abundant iguanids in the locality, rendering them good candidates for referral. (Certain other lizard taxa in the fauna are also large enough to have produced the vertebrae—even too large—but the clades to which these species generally lack accessory articulations and/or subcentral impressions. For a discussion of vertebrae in Squamata, see Hoffstetter and Gasc 1969). For the moment, I decline (semi-)formally to associate the vertebrae.
The most notable feature of Vertebra Type 1 is the development of zygosphene–zygantral accessory articulations. Zygosphenes and zygantra have evolved several times within Squamata, most notably in Iguania, Teiidae, and Serpentes (Hoffstetter and Gasc 1969; Estes et al. 1988). Within Iguanidae, they appear in all iguanines, some "Enyalioides," crotaphytines, and corytophanines (de Queiroz 1987) as well as some members of Tropidurinae* (e.g., Uranoscodon superciliosus) and Polychrotinae* (e.g., Anolis equestris, Polychrus). In lacking complete separation of zygosphene from prezygapophysis, Vertebral Type 1 has a condition intermediate between having no accessory articulations and having advanced ones (see de Queiroz 1987, figure 36), as in the extant iguanine Dipsosaurus dorsalis.
The squaring and pairing of the mid-sagittal ventral ridge is more restricted in distribution. I have observed it in some corytophanines, crotaphytines, some Sauromalus obesus, and Dipsosaurus dorsalis, especially on the more posterior presacral vertebrae; it appears to be absent in the large iguanines (including juveniles). The significance of this feature is uncertain.
![]()