 Referred specimens. PTRM 5359 (partial right dentary),
Figure 12; 5358 (right dentary fragment)
Referred specimens. PTRM 5359 (partial right dentary),
Figure 12; 5358 (right dentary fragment)
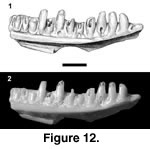
Description. Both specimens are small. PTRM 5359 is a partial right dentary with 13 full tooth positions and five relatively complete teeth (Figure 12). The IMS extends posteriorly to under the fifth tooth from the rear. A facet for the splenial, on the underside of the subdental shelf, extends to under the eighth tooth from the rear. There is a weak subdental shelf, but no sulcus dentalis. Two labial foramina are present, the posterior-most under the seventh tooth from the rear.
Tooth replacement is intradental ("iguanid" type of McDowell and Bogert 1954). The teeth, as preserved, are relatively blunt (especially in PTRM 5358, which is worn) and low-crowned. Many teeth are especially broad at the base, tapering toward their crowns. The crowns themselves show a moderate central cusp bordered by strong mesial and weaker distal crests that curve labially around the main cusp, forming "shoulders." The lingual, but not labial, surfaces of the crowns are striated.
Remarks. Scincoid MPH-1 is distinguished from heretofore described Paleogene scincoid lizards by the tapering of the teeth toward their apices, and the absence of a sulcus dentalis. The primary reason for allying it with Scincoidea is general tooth form, which, as described, consists of a main central cusp, striated lingually and bordered by "shoulders" (not distinct accessory cusps). Clearly, better specimens are desirable.
 Referred specimens. PTRM 2720 (partial left dentary),
Figure 13.1; 5360 (edentulous left dentary); 5361 (partial right dentary), Figure 13.2; 2074 (medial portion of right maxilla); 5284 (left dentary fragment); 5288 (partial right dentary); 2074 (right maxilla fragment); 2713 (partial right maxilla); 5298 (right maxilla fragment); 5387 (right dentary fragment)
Referred specimens. PTRM 2720 (partial left dentary),
Figure 13.1; 5360 (edentulous left dentary); 5361 (partial right dentary), Figure 13.2; 2074 (medial portion of right maxilla); 5284 (left dentary fragment); 5288 (partial right dentary); 2074 (right maxilla fragment); 2713 (partial right maxilla); 5298 (right maxilla fragment); 5387 (right dentary fragment)
Amended diagnosis. As in Holman (1972), except that the AIAF is located at or behind the ultimate dentary tooth in most members (see below) and tooth crowns are striated lingually.
Description. None of the Medicine Pole Hills specimens approaches the quality of the holotype spleniodentary of "Palaeoxantusia" borealis (SMNH 1435). The most complete specimen, PTRM 5360, is edentulous and broken just posterior to the ultimate tooth and lacks a splenial; it displays many features seen in the other specimens from the Medicine Pole Hills l.f. It is relatively small, with a tooth-row length of 4.0 mm; the height of the dentary at the ultimate tooth is 1.1 mm. A sulcus dentalis is well developed. Twelve tooth positions are present. The AIAF is situated just behind the ultimate tooth (more clearly seen in PTRM 2720: Figure 13.1). There are five labial foramina. PTRM 5361 shows the long, horizontal posterior extension of the dentary beyond the tooth row (Figure 13.2).
The teeth of most specimens are rather worn; they are blunt and unicuspid and do not preserve details of crown morphology. The teeth of a couple of specimens retain traces of the weak "shoulders" that lie mesial and distal to the central cusp. The ultimate maxillary tooth of PTRM 5298 and 5387 present clear examples of weak striations near the base of the crown.
Remarks. This Medicine Pole Hills xantusiid compares well to "Palaeoxantusia" borealis, and it is referred to that taxon. The specimens described here are smaller than those from the Calf Creek l.f. (Medicine Pole Hills: mean dentary depth at ultimate tooth ~1.1 mm, n = 4; Calf Creek: mean depth ~1.3 mm, n = 6). However, there is much overlap at the lower end of the Calf Creek size range, and a 20% difference in linear dimensions would translate into an approximately 75% difference in volume (and hence, mass), which accounts for the overall slender appearance of the Medicine Pole Hills specimens. It is possible that, if the specimens are correctly referred, "P." borealis exhibits an inverse correlation of size (mass) to latitude—and, hence, temperature—conforming to Bergmann's rule (Bergmann 1847), which is disputed in squamates (e.g., Angilletta et al. 2004). Alternatively, none of the specimens in the sample may be fully grown. Only one of the largest specimens from Calf Creek (depth ~1.5 mm) evinces complete fusion of the splenial to the dentary and can be interpreted as more-or-less fully adult. Unfortunately, none of the specimens preserves the splenial at all (this fact may indicate that the splenial was not yet fused, i.e., that all the specimens are from juveniles).
Fossil xantusiid species are typically based on the dentary or spleniodentary, and there are precious few features in this bone upon which phylogenetic hypotheses can be based (Vicario et al. 2003). Thus, it is important to point out a feature not yet described that may provide some constraint on the relationships of fossil xantusiids. In the earliest-known of these, "Palaeoxantusia" fera, from the middle Paleocene of Wyoming (Estes 1976; Sullivan 1982), the AIAF is located under the ultimate or penultimate dentary tooth in all specimens for which this character is clearly scorable (n = 4 and 9, respectively). The same holds true for the large Bridgerian form "P. fera" (YPM-PU 17506, n = 3) and the Bridgerian type of P. fera (AMNH 3815; Hecht 1956). The only specimen that appears to violate this generalization is AMNH 3821, from the same locality as the type of P. fera; it appears, then, that middle Eocene "Palaeoxantusia" exhibits some polymorphism. "Palaeoxantusia" kyrentos and "P." allisoni from the Uintan of California also display an anterior foramen (see Schatzinger 1980, text-figures 1,4). Among extant xantusiids, the anterior position of the AIAF is seen in Lepidophyma flavimaculatum (pers. obs.); some Xantusia riversiana (three of eight) also approach the condition. In all "Palaeoxantusia" borealis from both the Medicine Pole Hills (Figure 13.1) and Calf Creek, in contrast, the AIAF is located just behind the ultimate dentary tooth. This posterior position—it may be defined as the case in which the foramen is entirely located behind the midline of the ultimate tooth—is also seen in the living basal (Hedges and Bezy 1993; Vicario et al. 2003) member Cricosaura typica (see Savage 1963, figure 8), Xantusia downsi (see Norell 1989, figure 14), and X. vigilis, X. arizonae, X. henshawi, and a significant proportion of X. riversiana (pers. obs.). Given Vicario et al.'s (2003) recent phylogeny of Xantusiidae, it seems possible that the posterior position is primitive for Xantusia and partially reversed in X. riversiana. The only other small xantusiid, Cricosaura typica, must independently have acquired this feature, implying that it might simply be related to absolute size. On the other hand, the posterior position in one paratype of P. fera and many X. riversiana indicates that this need not be the case. Size correlation also suggests that an anterior shifting of the foramen ought to be visible during development.
Estes (1983:125) concluded that "Palaeoxantusia" borealis was inadequately diagnosed and might be synonymous with P. fera. The validity of "P." borealis is confirmed, and its diagnosis is amended above.
 Referred specimens. PTRM 1793 (right dentary),
Figure 14; 1825, 5364–5370, 5638, 5639, 5662, 5667, 5671, 5672, 5691 (trunk vertebrae)
Referred specimens. PTRM 1793 (right dentary),
Figure 14; 1825, 5364–5370, 5638, 5639, 5662, 5667, 5671, 5672, 5691 (trunk vertebrae)
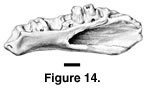
Description. The worn dentary is relatively large, the tooth row measuring 6.2 mm in length. There are eight tooth positions, but only the sixth and seventh teeth are even partially preserved (Figure 14). They are subpleurodont and slightly separated from the interior edge of the dentary by a weak subdental shelf. Their bases are inflated and confluent, rather square in coronal section. The first and fourth tooth bases are the largest. The splenial notch extends to the boundary between the fourth and fifth teeth. The interior of the dentary has a small ridge that extends ventrally from the top of the Meckelian canal; an intramandibular septum is therefore not well developed posteriorly. The Meckelian groove is closed and fused. There is a small, medially facing notch, just posterior to the last tooth, which is confluent with the well-defined coronoid fossa on the labial surface of the bone, thus excluding the possibility of a long labial process of the dentary. The coronoid fossa actually comprises two conjoined fossae, the boundary between which is marked by a faint ridge. The medial, shallower one is confluent with the aforementioned notch posterior to the last tooth; the lateral, deeper one extends anteriorly to the level of the boundary between the sixth and seventh teeth. The coronoid fossa as a whole is triangular in shape, with the apex pointed forward. Two, widely spaced labial foramina are present, one each below the second and fifth teeth. The mandibular symphysis is weak.
The best-preserved vertebra (PTRM 5364) is large, measuring 7.5 mm in length (along neural arch) and somewhat wider anteriorly than posteriorly (10.2 mm at prezygapophyses vs. 9.1 mm at postzygapophyses). The prezygapophyses are oriented in a nearly coronal plane. A midline keel runs along the entirety of the neural arch; the keel ends in a projection (partially broken) from the posterior end of the arch. The posterior end is flared and markedly taller than the anterior end. On the dorsal surface of the posterior half of the neural arch run linear ridges (or "flutings": Gans 1978) which diverge posteriorly from the midline. The ridges match corresponding structures on the ventral surface of the arch (a convexity above matches a concavity below), and thus, viewed from behind, the edge of the arch appears crinkled. (This ventral mirroring of flutings is present in many, but not all, vertebrae referred to cf. Rhineura sp.) The centrum is depressed. The condyle has a constricted neck. There is no hypapophyseal ridge; only the right subcentral arterial foramen is present, and it is located close to the synapophysis, well away from the midline. The synapophyses are hemispherical. The vertebrae (including PTRM 5364) typically have a small indentation just above the synapophysis on their lateral side. Most referred vertebrae are only two-thirds the size of PTRM 5364.
Remarks. The low tooth count and subpleurodont tooth implantation are sufficient to refer this species to Amphisbaenia. Unfortunately, the polarity of many character state changes seen within Amphisbaenia is equivocal because of uncertain in-group and out-group relationships (Estes et al. 1988; Kearney 2003); this complicates discussion of relationships. The determination of whether many features are primitive or derived in Rhineuridae depends crucially on the position that that taxon occupies. Despite these phylogenetic difficulties, a plethora of rhineurid amphisbaenians has been named from the late Eocene through Oligocene of North America, generally based on more-or-less complete skulls. PTRM 1793 shares with many of these fossil rhineurids—including the middle Eocene Spathorhynchus fossorium Berman (1973), early Oligocene Rhineura hatcheri (Kearney et al. 2005), Miocene Rhineura sepultura Holman (1979), and extant R. floridana (Cope 1892, figure 5; pers. obs.)—enlargement of the first and fourth dentary teeth. In "Rhineuridae sp. indet." from the late Paleocene of Wyoming (Estes 1975), only the fourth tooth appears to be enlarged (pers. obs.). Hyporhina tertia has enlarged first and fourth "maxillary teeth" (Berman 1972: 8); whether this applies to the dentary is unclear. The first and fourth dentary teeth appear to be enlarged in Jepsibaena minor (YPM-PU 13460; pers. obs.). It is probable that specified tooth enlargement is diagnostic of a clade including part of the stem of R. floridana, though precisely how much of the stem is at this point difficult to assess. The late Paleocene "Rhineuridae sp. indet." is the earliest taxon to display apomorphy one (fourth tooth enlarged); apomorphy two (first tooth enlarged) is first known in the early Eocene J. minor. Kearney (2003) concluded that all these North American forms were more closely related to R. floridana than to any other living amphisbaenian.
Cf. Spathorhynchus sp. can be excluded from described species of Rhineura by the primitive retention of eight dentary teeth. Spathorhynchus fossorium and the late Eocene S. natronicus also retain the presumably primitive state of eight dentary teeth (Berman 1973, 1977) and are both relatively large, with tooth row lengths somewhat less than 10 mm. Thus, of described coeval rhineurids, PTRM 1793 compares best to Spathorhynchus natronicus; due to poor preservation, however, I refer the specimens to cf. Spathorhynchus sp.
The largest vertebra, PTRM 5364, is considerably larger than one would expect in relying on the dentary alone. This fact suggests that the animal represented by PTRM 1793 was not yet fully grown. Cf. Spathorhynchus sp. may be the same as the large rhineurid taxon described as Lestophis sp. nr. L. anceps by Holman (1972) based on vertebrae. [Estes (1983), with good cause, declared Lestophis a nomen dubium.] Berman (1972) stated that ridges are present on the neural arches of vertebrae in rhineurids, to which list Estes (1976) added Amphisbaena (in A. alba, this is particularly true of the more anterior vertebrae). Kearney (2003) recently found dorsal ridges to be present also in Aulura, Dalophia, Leposternon, and Monopeltis, which she regarded as basal in Rhineuroidea. Flutings are not always reflected on the underside of the neural arch, but this is conceivably related to size.
Interestingly, amphisbaenian vertebrae—from both cf. Spathorhynchus sp. and cf. Rhineura sp. (see below)—appear to be unusually abundant in the Medicine Pole Hills l.f. Iguanids, though they are much more diverse and numerically abundant (based on jaw elements), are represented by (relatively) far fewer vertebrae. The same might be true of the amphisbaenians in the Calf Creek l.f., which are represented by 58 and 43 vertebrae (small and large, respectively; Holman 1972). Even considering that a given amphisbaenian typically has more than three times as many vertebrae as a non-fossorial squamate (<30 precloacal vertebrae in non-fossorial taxa versus often >100 in amphisbaenians; Hoffstetter and Gasc 1969), the difference appears anomalous.
 Referred specimens. PTRM 5248 (left dentary),
Figure 15; 5322 (right dentary fragment); 5725 (left dentary); 1422, 1434, 1438, 1443–1445, 1553, 1796, 1812, 1813, 1828, 1831, 1851, 2023, 2042, 2067, 5171, 5180, 5216, 5235, 5238, 5348 (12 specimens), 5362, 5363, 5384–5386, 5668, 5673, 5674, 5686–5688 (trunk vertebrae)
Referred specimens. PTRM 5248 (left dentary),
Figure 15; 5322 (right dentary fragment); 5725 (left dentary); 1422, 1434, 1438, 1443–1445, 1553, 1796, 1812, 1813, 1828, 1831, 1851, 2023, 2042, 2067, 5171, 5180, 5216, 5235, 5238, 5348 (12 specimens), 5362, 5363, 5384–5386, 5668, 5673, 5674, 5686–5688 (trunk vertebrae)
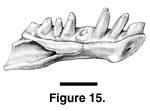
Description. The most complete specimen, PTRM 5248, has a tooth row length of 3.0 mm. There are seven tooth positions (Figure 15), although the fourth tooth is missing. Of these seven, the first and fourth are enlarged relative to the others. The teeth are subpleurodont and conical, tapering uniformly toward their crowns; these crowns, preserved only on the third and fifth teeth, are slightly recurved. There is a narrow, shallow sulcus dentalis. The symphysis is relatively large, expanded relative to the part just distal to it and has a small posteroventral projection. The Meckelian groove is closed and fused, and the splenial notch extends anteriorly to the level of the boundary between the fifth and sixth teeth. A fossa for the labial coronoid blade extends anteriorly to the level of the seventh (last) tooth.
PTRM 5322 is approximately the same size as PTRM 5248, though it is broken (just?) in front of the anterior margin of the splenial notch and preserves only the first through fifth teeth. The first and fourth teeth are enlarged. The symphysis is relatively large, and it has a posteroventral projection as in PTRM 5248.
The vertebrae of cf. Rhineura sp. are quite abundant and typical of rhineurid vertebrae. Vertebral flutings (see above under cf. Spathorhynchus sp.) are present but are only rarely reflected on the underside of the neural arch (e.g., in PTRM 5171), and then only weakly. The subcentral arterial foramina are typically located along the anterior third to fourth of the centrum. As in cf. Spathorhynchus sp., there is typically an indentation dorsal to the synapophysis on the lateral side of the vertebrae.
Remarks. The referral of these specimens to cf. Rhineura sp. is based on small size, tooth count, and age. The reduction to seven dentary teeth is a derived feature within Rhineuridae (see above). Cf. Rhineura sp. is therefore closer to Rhineura than are various Paleogene forms like Jepsibaena minor (8 dentary teeth) and Spathorhynchus spp. (8). No known North American Eocene rhineurid has seven dentary teeth, but Gilmore (1928) describes early Oligocene Rhineura hatcheri (CM 423A and B) as having seven. Hyporhina antiqua (YPM-PU 11390) also has seven dentary teeth, but that species lacks the expanded symphysis seen in J. minor and cf. Spathorhynchus sp. (pers. obs.) The enlargement of the first and fourth teeth places cf. Rhineura sp. firmly on the rhineurid line, and its possession of only seven dentary teeth indicates that it is derived relative to Spathorhynchus.
Sullivan and Holman (1996) synonymized many early Oligocene rhineurid taxa, arguing that the characters used to distinguish fossil amphisbaenians, including tooth count, may be related solely to ontogenetic stage and therefore "taxonomically insignificant" (p. 362). This point is well taken, for it is widely recognized, for instance, that tooth count increases with absolute size during development in many squamates. (I am not aware of any such study specifically on amphisbaenians.) Estes (1983) was correct in stating that taxonomic and systematic revision of Paleogene amphisbaenians is needed.
Neverthelesss, the dismissal of tooth count data in rhineurid systematics seems not yet to be warranted. Granted, extant Rhineura floridana has only six dentary teeth, and it is also smaller than many of its presumed early Paleogene relatives, which have eight dentary teeth. But Rhineura floridana's small size is itself an evolutionary change. Its small size and low tooth count may be informative for both alpha taxonomy and systematics, even if the two characters are developmentally correlated and might be considered a single character complex. In other words, ontogenetic correlation is by itself an insufficient basis for discarding character information in the referral of fossil forms. Additionally, in the particular case of Rhineuridae, absolute size is clearly a poor predictor of tooth count: the large Eocene Spathorhynchus as well as small Jepsibaena minor have eight dentary teeth, whereas the large Miocene Macrorhineura and small extant Rhineura floridana have but six.
Cf. Rhineura sp. might be the same as the small rhineurid species noted by Holman (1972) in the Calf Creek l.f., which was represented only by vertebrae.
 Referred specimens. PTRM 5375 (partial left dentary),
Figure 16; PTRM 1824 (partial right dentary); PTRM 5376 (anterior left dentary fragment)
Referred specimens. PTRM 5375 (partial left dentary),
Figure 16; PTRM 1824 (partial right dentary); PTRM 5376 (anterior left dentary fragment)
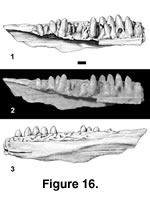
Description. The most complete specimen of this taxon is PTRM 5375. It is large, the tooth row being 13.7 mm long as preserved. A weak, narrow sulcus dentalis (Figure 16.1–2) is present at the anterior end of the bone. Fourteen or fifteen tooth positions are preserved. The teeth are pleurodont and low-crowned, less than one-third of their height projecting above the parapet of the jaw. The tooth crowns are striated lingually, and small mesial and distolabial crests descend the crowns (the mesial crest is much stronger), creating a V-shaped cutting edge. Except for the most posterior teeth, the crowns are discolored and somewhat transparent. They are bluntly pointed (in part due to wear), slightly recurved, weakly labiolingually compressed, and a little concave lingually near the juncture between crown and shaft. The teeth become progressively smaller and more blunt posteriorly and lose the crown-shaft concavity. The excavated base of the third preserved tooth with the incoming replacement tooth not directly below it indicates an interdental mode of replacement. The Meckelian groove is open. Though much of the shelf that supports the teeth has been broken posteriorly, a thin upper remnant of the IMS is evident; its original extent is unknown.
Externally, labial foramina are not readily identifiable in PTRM 5375, but a groove runs along much of the length of the dentary in the expected position of these foramina (Figure 16.3). The posterior, ventrolateral surface of the dentary is flattened, or planated; the plane of flattening is parallel to the longitudinal axis and oblique to a sagittal plane. Articulation surfaces with the postdentary bones are not apparent.
The two other specimens are far less complete, but both of them confirm the tooth morphology of this taxon. Additionally, PTRM 1824 displays the planated, external dentary surface and labial groove in the position of the labial foramina. There appear to be a couple of foramina present along the groove; whatever their number, they are small. Its two remaining teeth also exhibit discoloration of the crowns. PTRM 5376 exhibits a weak anterior sulcus dentalis but does not preserve the portion of the dentary that is planated in the other two specimens. No discoloration of the tooth crowns is evident in this specimen.
Remarks. The interdental mode of tooth replacement unites this new species with Anguimorpha (cf. Estes et al. 1988). Its tooth morphology is unique, but the V-shaped cutting edge is found in Anguidae and some anterior teeth in Xenosaurus (Gauthier, 1982; pers. obs.). In possessing lingually striated tooth crowns it also likens anguids, Xenosaurus, and many Varanus (pers. obs.). Many other features, such as osteodermal characteristics and the state of the IMS posteriorly, cannot yet be evaluated. Oddly, nearly all anguimorphs possess at least a moderate subdental shelf along most of the dentary. That this shelf is essentially lacking in Anguimorph MPH-1 would seem to be autapomorphic.
Relationships among anguimorphs generally and anguid subclades in particular are not well resolved (e.g., Camp 1923; Rieppel 1980; Gauthier 1982; Macey et al. 1999; Wiens and Slingluff 2001; Townsend et al. 2004), which also limits our understanding of character evolution within the group. Preserved features in Anguimorph MPH-1 appear only to disbar it from membership in particular anguid subclades: from Diploglossinae and Gerrhonotinae, by its lack of bicuspid teeth; from Glyptosaurinae by its tooth morphology; and from Anniella, by its higher tooth count and tooth morphology. In all these respects, Anguimorph MPH-1 is primitive relative to known representatives of these groups. Although its tooth form and lack of a significant subdental shelf render Anguimorph MPH-1 distinctive, better specimens are required before a formal new name can be applied.
 Referred specimens. PTRM 1807 (left dentary fragment),
Figure 17.1; 1823 (jaw fragment); 5378 (osteoderm),
Figure 17.2
Referred specimens. PTRM 1807 (left dentary fragment),
Figure 17.1; 1823 (jaw fragment); 5378 (osteoderm),
Figure 17.2
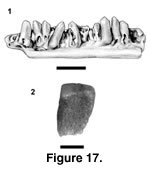
Description. PTRM 1823 is a fragment of either a right dentary or a left maxilla. The bit of bone to which the two preserved teeth are attached is worn. The crown of the first tooth is also worn, but the second does not project as high—it is probably an incompletely emplaced replacement tooth—and appears largely undamaged. This second (posterior) tooth bears a distinct mesial crest, offset from the rest of the tooth crown by a shallow groove. The main tooth cusp is distally displaced and slightly labiolingually compressed. The lingual surface of the crown is smooth, but near the base of the crown, on its mesial side, there are coarse striations; the striae do not extend onto the lingual side, possibly a result of wear. The bases of both teeth are missing, so it is difficult to estimate crown height.
There are 13 tooth positions, with eight teeth, preserved in PTRM 1807, a left dentary fragment (Figure 17.1). The teeth are considerably smaller than in PTRM 1823. They are of moderate height, and the lingual surfaces of the crowns are apparently unstriated. All but the anterior-most teeth have strong mesial crests; on posterior teeth these crests are so expanded that the crowns are mesiodistally longer than the shafts. A few teeth appear fully implanted, their labial margins flush with the parapet of the jaw; lingual to these teeth is a moderate subdental shelf. Many of the preserved teeth are incompletely emplaced replacement teeth and have bulbous, rounded bases that are not strongly fused to the dentary; they also sit away from the parapet of the jaw over the subdental shelf. The relative position of replacement teeth is difficult to determine posteriorly, where the tooth pattern is distorted. The anterior-most tooth, however, is excavated distolingually, indicating an interdental (anguimorph) pattern of tooth replacement.
The osteoderm, PTRM 5378 (Figure 17.2), is rectangular and unkeeled, ~2.2 mm in length. Its sculpturing consists of variably linked, apically flattened tesserae that are separated by grooves; where they merge, they create a vermiculate sculpture. The anterodorsal edge of the gliding surface was oblique to the anterior margin, but there is no indication of a posterior projection of the gliding surface. This surface is approximately one-fourth the length of the whole osteoderm.
Remarks. The jaw fragments constitute the primary evidence for the presence of a diploglossine in this assemblage. Strong development of the mesial tooth crests, often with the development of a secondary cusp, diagnoses Diploglossinae and possibly Gerrhonotinae (within Anguidae) as well as Xenosaurus and its stem (Gauthier 1982); only diploglossine anguids have apomorphically shorter, stocky teeth, often with expanded crowns that are mesiodistally longer than their shafts. An undamaged osteoderm with a posterior projection of the gliding surface would be desirable to buttress this conclusion (cf. Hoffstetter 1962; Gauthier 1982). The sculpturing of the osteoderm, however, is similar to that of the (stem) diploglossine Eodiploglossus borealis Gauthier (1982) from the early Eocene of Wyoming; and in living diploglossines and E. borealis, not all the osteoderms have a posterior projection. There is presently little that would permit a more specific determination of the taxon, nor is it known whether it lies on the stem of Diploglossinae or in the crown. It represents the latest-known occurrence of the diploglossine lineage in central North America.
Referred specimens. PTRM 2606 (partial left dentary); PTRM 2617 (partial right maxilla); 5272 (jaw fragment); 5381 (right maxilla); 5685 (right frontal fragment)
Description. PTRM 2606 is the anterior two-thirds of a left dentary. Fourteen tooth positions and two teeth (numbers 4 and 10, from symphysis) are preserved. Both teeth, like the dentary itself, are streamworn, and details of crown morphology are lacking. The apex of tooth 10, however, is somewhat labiolingually compressed. The crowns are moderate in height, projecting less than 50% above the parapet of the jaw. There is a moderately developed subdental shelf, without attendant sulcus dentalis; this shelf is largely broken anteriorly. The Meckelian groove is ventrally located on the anterior portion of the dentary. There are six, irregularly spaced labial foramina on the external surface. There is also a small indentation at the posterolateral margin of the preserved portion, but the generally streamworn character of the specimen suggests that this feature may represent a rounded fracture rather than the anterior edge of an articular surface or fossa.
PTRM 5381 is a small, nearly complete but somewhat worn right maxilla. The premaxillary process is broken near the base of the nasal process. Eleven tooth positions and seven complete or partial teeth are preserved. The teeth are widely spaced, but it is impossible to estimate crown height. Both the crowns and the shafts are somewhat recurved. The crowns are unicuspid, blunt, and without labial or lingual striations, which may or may not be due to abrasion. They decrease in height posteriorly such that the posterior-most teeth are exceedingly small. The dorsal surface of the maxilla is divided into anterior and posterior segments by an oblique ridge on the interior surface of the nasal process and dorsal surface of the main body of the maxilla. The SAF is located at the level of the posterior edge of the nasal process. The foramen is set in a short groove and is covered anteriorly by the dorsal roof of the maxilla. The palatine process is only weakly developed and is located at about the same level as the superior alveolar foramen; on its dorsal surface is a shallow facet for articulation with the palatine. Laterally, there are four labial foramina and several other foramina, irregularly spaced, on the slightly irregular surface of the nasal process. There are no osteoderms fused to the surface of the nasal process, probably a result of the small size of the individual. Another poorly preserved maxilla (PTRM 2617) appears to have weak striations on the lingual surface of one anterior tooth.
The frontal fragment (PTRM 5685) is heavily worn, but it certifies the existence of a non-glyptosaurin glyptosaurine in the Medicine Pole Hills l.f. A thin strip along the orbital margin is devoid of the tuberculated osteodermal crust that otherwise covers the bone. A small foramen pierces the lateral margin of the orbit.
Remarks. These specimens are compared to Peltosaurus on the basis of glyptosaurine tooth form, size, and geologic age. PTRM 2606 shares with specimens of Peltosaurus granulosus robust teeth, similar tooth spacing, and similar size, but more complete teeth, as on the maxilla, seem rather more recurved than in P. granulosus. The lack of striations on most teeth may be attributable to stream abrasion, a phenomenon that may also account for the lack of striations on Peltosaurus sp. from the Cypress Hills Formation of Saskatchewan (see Lambe 1908; cf. Estes 1983). Peltosaurus is rare in late Eocene sediments, though one would expect a "melanosaurin" glyptosaurine to be present somewhere on stratigraphic grounds.
 Referred specimens. PTRM 1825 (left parietal fragment):
Figure 18; 5647, 5709 (cephalic osteoderms)
Referred specimens. PTRM 1825 (left parietal fragment):
Figure 18; 5647, 5709 (cephalic osteoderms)
Description. PTRM 1825 is a left, anterior fragment of parietal. Its edges and surfaces are in places smoothed by abrasion, including the tops of the osteoderms (Figure 18.1). There are four relatively complete, tall, bulbous, tuberculated osteoderms coössified with the dorsal surface. The osteoderms are vaguely polygonal, with subrounded corners, and separated from one another by deep grooves. They are undercut in places, and some isolated tubercles occur in the grooves. Several osteoderms are missing from the anterolateral part of the bone. The left edge of the parietal foramen (Figure 18.1–2, pf) is visible, perforating the first, large, median osteoderm. In medial view, the parietal fragment has a hemispherical cavity, about 2.5 mm in diameter, that excavates both osteoderm and underlying bone. It opens dorsally through the osteoderm and ventrally through a short canal into the cranial cavity. On the ventral surface of the parietal, the anterior ridges (Figure 18.2) are well developed. The distance between the median plane and the anterior ridge is less than that between the anterior ridge and the lateral edge of the parietal (i.e., the triangular inter-ridge area is small). There are two notches, rounded at their medial apices, on the anterolateral part of the ventral surface; they are confluent with the anterolateral edge of the bone and constitute articulation facets for the postfrontal and possibly also the postorbital.
PTRM 5647 and 5709, hexagonal cephalic osteoderms, are also referred to this taxon.
Remarks. The more-or-less hexagonal osteoderms of PTRM 1825 are sufficient to refer it to Glyptosaurini. As Sullivan (1979) noted, tall, bulbous osteoderms, among other features, serve to distinguish the presently monotypic Helodermoides tuberculatus from other glyptosaurins. Sullivan also argued (op. cit.) that H. tuberculatus is diagnosed by "closure of the supratemporal fenestra," a phenomenon that in Glyptosaurini deserves further investigation. The primitive condition is illustrated by Paraglyptosaurus princeps ("Glyptosaurus hillsi": Gilmore 1928, figure 63; and "G. rugosus": Meszoely 1970, figure 16B; see Sullivan 1979 for nomenclature), in which the postfrontal articulation on the parietal is a single notch restricted to the latter bone's anterior-most end. Sullivan (1979, figure 10h) figures complete closure of the supratemporal fenestra in one specimen of Helodermoides (USNM 214736). The condition in PTRM 1825 is intermediate, for although the two notches on the ventral surface suggest some closure, the bone thins out laterally, and there are no other obvious articulation surfaces. Sullivan is surely correct that extensive closure of the supratemporal fenestra by increased articulation of the temporal bones is a derived feature in Anguidae. The seemingly significant difference in supratemporal closure between specimens (PTRM 1825 and AMNH 8706 vs. USNM 214736) suggests specific difference. However, because the type specimen of H. tuberculatus (CM 707) consists only of frontals, a determination of which form is new is not yet possible. Helodermoides is a previously known and in this sense unproblematic taxon from the Chadronian.
 Referred specimens. PTRM 1986 (premaxilla),
Figure 19; 5371 (dermal skull bone fragment)
Referred specimens. PTRM 1986 (premaxilla),
Figure 19; 5371 (dermal skull bone fragment)
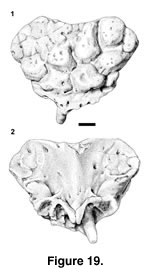
Description. PTRM 1986 is a slightly worn but relatively complete premaxilla. The dorsal surface is covered by coössified osteoderms, worn on their outer surfaces (Figure 19.1). The osteoderms are broad and rounded and separated by narrow valleys. They tend to be flatter toward the midline and more conical laterally and do not extend to the extreme lateral margins of the bone. Tiny pocks and perhaps foramina pierce them. The anteroventral margin of the bone, below the fused osteoderms, is perforated by several small foramina. The posterolateral borders of the premaxilla appear unbroken. The dorsal nasal process is broken near its tip. On the ventral margin, there are spaces for seven teeth, including one midline tooth (Figure 19.2). Only one tooth, the first, right, paired tooth, is preserved, and it is short, the crown worn and blunt. The incisive process is narrow and bilobed, unfused even basally, where the right and left portions are strongly apposed; it lies immediately behind the median tooth alveolus. The ventral premaxillary foramina are located lateral to the incisive process and posterior to the first pair of premaxillary tooth alveoli. The posterior premaxillary foramina are larger than the ventral ones, with which they connect internally (Bahl 1937). Broad dorsal wings extend from the midline to the lateral edge of the bone, covering the typical location of the external nares. The wings are thinner between the thick midline and the thick lateral margins. Small grooves present on both sides of the ventral surface mark articulations with the premaxillary processes of the maxillae.
PTRM 5371 is a thin, rectangular fragment of a skull bone, 5.5 x 2.5 mm. It is referred to the same taxon as the premaxilla on the basis of size and similar osteoderms. All of the osteoderms are flatter than on the premaxilla, however: none is bulbous. One long edge is unbroken and is grooved for an articulation along the whole of its preserved length. In internal (ventral?) aspect, this same edge is distinguished by a moderately high ridge. Its appearance suggests that the element may bound the supratemporal fenestra (a parietal fragment, or postfrontal), which might have been partially closed.
Remarks. These specimens are referred to an undetermined xenosaurid with some hestitation. In possessing multiple anterior foramina the premaxilla likens Xenosaurus but differs from most other anguimorphs. But unlike Xenosaurus platyceps and X. grandis, which have nine premaxillary teeth (like most anguimorphs), Xenosaurid MPH-1 has but seven. Uncertain phylogenetic relationships within Anguimorpha, however, and the presence of only seven premaxillary teeth in Shinisaurus crocodilurus, render it uncertain whether nine teeth in Xenosaurus is primitive or derived. Gilmore (1941) reported only eight in Exostinus serratus.
An alternative hypothesis is that this species is a helodermatid, which would be consistent with the more bulbous character of the osteoderms on the premaxilla. The absence of the "coralline" sculpturing characteristic of Heloderma might then be interpreted as artifactual (i.e., it has been removed by abrasion). The premaxilla is too small (especially considering the fusion of the osteoderms) to be attributed to Helodermoides and moreover lacks any indication of tubercles. There is presently no evidence that would tie the specimens to any other species in the fauna.
Regardless of its affinities, the broad posterolateral wings of the premaxilla of Xenosaurid MPH-1 are unlike anything known in squamates. The external nares must have been displaced a considerable distance posteriorly as a result of their presence; furthermore, the maxilla necessarily articulates on the ventral surface of the premaxilla rather than the dorsal surface, as in most lizards.
 Holotype. PTRM 5379 (partial left dentary),
Figure 20.1
Holotype. PTRM 5379 (partial left dentary),
Figure 20.1
Paratype. PTRM 5380 (partial parietal), Figure 20.2–3
Locality and horizon. PTRM V89002, Chalky Buttes Member, Chadron Formation (late Eocene), North Dakota, USA
Distribution. Thus far only known from type locality.
Etymology. L. eduro, meaning "to last, endure, persist," in reference to its status as the last known varanid in North America
Diagnosis. A varanid lizard with teeth that are laterally compressed somewhat obliquely to the jaw axis and have strongly recurved tooth crowns, ventromedially expanded tooth bases, and coarse if discontinuously expressed infoldings along the tooth base labially and lingually; dentary with a wide subdental shelf posteriorly; frontoparietal suture weakly anteriorly convex; and dorsal surface of parietal highly rugose.
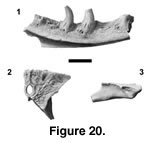
Description of holotype. PTRM 5379 is the medial portion of a large left dentary, preserving six tooth positions with three partial to near-complete teeth (Figure 20.1). The bases of the teeth are expanded posteromedially. They show coarse but discontinuously expressed infoldings; they are visible externally at the very base and then further up the shaft, but are subdued in between. Each tooth has one small, basal nutrient foramen, located distal to the anteroposterior mid-point of the tooth. The crowns lack serrations and are strongly recurved and labiolingually compressed, giving rise to mesial and distal carinae. The best-preserved tooth is the fourth, which shows a peculiar crown morphology. In the upper crown of this tooth, the plane of compression is not quite parallel to the dentary; rather, the anterior edge is rotated labially ~20°. A rigorous tooth count is unobtainable owing to incomplete preservation; by comparison with Varanus, the decreasing size of the posterior-most tooth bases and the rise of the subdental shelf indicate that the last preserved tooth is close to or at the end of the tooth row, and the narrowing of the Meckelian groove anteriorly suggests that probably 5–7 more teeth were present there. Tooth count may not have differed much from extant Varanus. Posteriorly, the subdental shelf is wide, about equal to the width of the tooth bases there. The Meckelian groove is in a ventral position anteriorly, and there is no indication how far anteriorly the splenial extended. The IMS extends posteriorly to the end of the preserved part of the dentary. The posterior, dorsolateral surface is strongly beveled, presumably just anterior to where the coronoid fit. Several labial foramina are present, the posterior-most being the largest.
Description of paratype. PTRM 5380 is the well-preserved, right side of a large, azygous parietal, referred to the same species on the basis of size and varanid morphology. The anterior margin of the bone—the frontoparietal suture—is weakly convex. Its dorsal surface is distinctly rugose, perhaps reflecting a strong attachment of epidermal scales (Figure 20.2). On the anterior portion of the bone, rugosities are bounded by widely spaced grooves, perhaps indicating the presence of relatively large scales. Posterolateral to the parietal foramen, tuberculae are smaller and more clearly developed (Figure 20.2) but probably smaller than individual epidermal scales. The parietal foramen is surrounded by a raised ridge nearly hexagonal in shape. The foramen was probably covered by a single, hexagonal interparietal scale and is relatively large (1.9 x 1.1 mm). A weak, longitudinal ridge extends anteriorly from the ridge surrounding the foramen to the frontoparietal suture (where it may have continued onto the frontal). Posteriorly, the jaw adductor musculature originates in the supratemporal fossa, whereas anteriorly the parietal table overhangs the fossa. On the anterolateral surface of the bone is a triangular notch for the attachment of the postfrontal (Figure 20.3); its limit is marked by a bump in the margin of the parietal table in dorsal view (Figure 20.2). Along the edge of ventral crista runs a small groove.
Remarks. Saniwa edura was probably close in size to, or slightly larger than, Saniwa ensidens Leidy (1870), the most well-known North American fossil varanid. The dentary of S. edura is slightly larger than that of the type specimen of S. ensidens: Gilmore (1928) notes that in the holotype of S. ensidens, three dentary teeth take up a total of 11.0 mm, whereas in PTRM 5379, three middle dentary teeth take up ~12 mm. There are at least three partial parietals of Saniwa* known [YPM 1074 (illustrated in Gilmore 1928, plate 9, figure 11), YPM 1059 (mentioned by Gilmore), and YPM 613], which Estes (1983) suggested were referable to S. ensidens. PTRM 5380 is smaller than them and clearly belonged to an animal smaller than the one that produced the dentary.
The dentary of Saniwa edura is similar to that of S. ensidens in having a wide subdental shelf posteriorly. In contrast, in living Varanus (except V. komodoensis) the shelf is much narrrower toward the posterior end of the dentary. The parietal of Saniwa edura distinguishes it from Saniwa ensidens. In YPM 1074 and 613, the lateral edges of the parietal table are sharp and upturned (Gilmore 1928), whereas in PTRM 5380 they are flat. This feature is virtually unknown in extant Varanus (it was only observed in V. niloticus and large V. exanthematicus, which have blunt, upturned edges). Additionally, the parietal table is rugose in S. edura, whereas it is smooth in S. ensidens and most Varanus (less well-developed rugosities of similar form are seen in V. prasinus). Comparison of tooth morphology is difficult owing to incomplete preservation of the type material of S. ensidens (Caldwell 2003).
Some aspects of the anatomy of Saniwa edura are rendered uncertain due to its poor preservation. For example, some modern species (e.g., Varanus melinus) and Palaeosaniwa canadensis have serrations on the mesial and distal tooth carinae; it is possible that such were present on PTRM 5379 but have been lost to stream abrasion. Similarly, many living Varanus show distinct lingual striations on the tooth crown; these were not observed in the holotype of S. edura. Several other features are primitive. For instance, the narrow grooves, extending anteriorly along ventrolateral edges of the parietal, were found in all varanoids examined and in extant taxa appear to correspond to the taeniae marginales of the synotic tectum. Likewise, the dorsal origin of the adductor musculature is shared with Lanthanotus and Varanus (Estes et al. 1988) together with fossil "necrosaurids" (Estes 1983).
Several medium-sized varanid species have been described from the middle Eocene, all now assigned to Saniwa* (see Gilmore 1928). The new species is also assigned to that taxon on the basis of overall similarity. Although Saniwa* presently has no known synapomorphies (Estes 1983), neither can it be shown to be paraphyletic. Assignment of edura to Saniwa* is a conservative solution: should new evidence later show Saniwa* to be paraphyletic, edura can always be removed. These specimens constitute the latest certain record of Varanidae in North America (cf. Estes 1983; Sullivan and Holman 1996). Brattstrom (1955) and Schatzinger (1975) have described Saniwa brooksi from the Uintan of California on the basis of vertebrae and a jaw fragment. Emry (1973) reported Saniwa sp. and ?Thinosaurus (= Saniwa) sp. from the White River Formation (Chadronian) of Wyoming but did not describe or cite specimens.
![]()