cf. Aciprion sp.
Iguanid MPH-3
Iguanid MPH-4
Cypressaurus Holman (1972)
Cypressaurus sp.
Autarchoglossa Wagler (1830)
Scincomorpha Camp (1923)
Scincoidea Oppel (1811)
Scincoid MPH-1
Xantusiidae Baird (1858)
Palaeoxantusia Hecht (1956)
"Palaeoxantusia" borealis Holman (1972)
Amphisbaenia Gray (1844)
Rhineuridae (Vanzolini 1951)
Spathorhynchus Berman (1973)
cf. Spathorhynchus sp.
Rhineura Cope (1861)
cf. Rhineura sp.
Anguimorpha Fürbringer (1900)
Anguimorph MPH-1
Anguidae Gray (1825)
Diploglossinae Cope (1864)
Diploglossine MPH-1
Glyptosaurinae (Marsh 1872)
Peltosaurus Cope (1872)
cf. Peltosaurus sp.
Helodermoides Douglass (1903)
Helodermoides sp.nov.?
?Xenosauridae Cope (1866)
Xenosaurid MPH-1
Varanidae Gray (1827)
Saniwa* Leidy (1870)
Saniwa edura sp. nov.
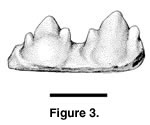 Referred specimens. PTRM
2038 (jaw fragment), Figure 3
Referred specimens. PTRM
2038 (jaw fragment), Figure 3
Description. A thorough description of this specimen was provided by Pearson (1998), who wrote:
The referred specimen PTRM-2038 is a fragment of bone ... with two tricuspid, acrodont teeth. The anterior tooth measures 0.87 mm in width and the posterior tooth 1.06 mm, as measured at their widest point. The teeth are laterally compressed with three anteroposteriorly aligned cusps, the median being the tallest at 0.95 mm for the anterior and 1.12 mm for the posterior tooth. The teeth are separated with a spacing of 0.27 mm at the narrowest point and 0.35 mm at the level of the jaw parapet. The total space occupied by both teeth ... equals 2.07 mm. The anterior tooth extends 0.50 mm above the parapet of the jaw and the posterior tooth 0.64 mm. Both teeth exhibit sub-equal lateral cusps.... The bases of the teeth are broad and fused to the dorsal margin of the [jaw] at a slight angle. ... The presence of lateral cusps on both teeth indicates the specimen originated from the middle, or more posterior portion, of the [jaw]. (p. 37)The specimen is illustrated in Figure 3.
Remarks. Tinosaurus* is a poorly diagnosed taxon erected for small- to medium-sized Paleogene squamates that evince acrodont tooth implantation and tricuspid posterior teeth (Estes 1983). It represents one or more incursions of acrodont iguanians into North America during a time of widespread faunal interchange (Gauthier 1982) that resulted from decreased latitudinal temperature gradients (Greenwood and Wing 1995). The type specimen (now at USNM) of the type species (T. stenodon Marsh 1872) is from the Bridgerian of North America. Barring multiple invasions by Acrodonta, the North American taxa may constitute an exclusively monophyletic group (though this cannot be demonstrated with apomorphies at present). The relations of these taxa to Asian and European species referred to Tinosaurus* are also unclear, but the North American taxa would remain the name-bearers.
Pearson (1998) compared PTRM 2038 to T. stenodon. There is presently not enough material to justify specific assignment. This specimen is the latest known record of Acrodonta in the New World (Pearson 1998).2
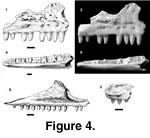 Holotype. PTRM 1841 (partial right maxilla),
Figure 4.1–2
Holotype. PTRM 1841 (partial right maxilla),
Figure 4.1–2
Paratypes. PTRM 2637 (right dentary fragment); 1990 (partial right dentary); 2596 (left dentary fragment); 1418 (right dentary fragment); 2673 (right dentary fragment); 5351 (edentulous right dentary), Figure 4.4–5; 5400 (jaw fragment); 1997 (right maxilla fragment); 5372 (left maxilla fragment); 5373 (right maxilla fragment), Figure 4.3; 5374 (partial right maxilla)
Locality and horizon. PTRM V89002, Chalky Buttes Member, Chadron Formation (late Eocene), North Dakota, USA
Distribution. Thus far known only from type locality.
Diagnosis. A polychrotine iguanid with a dorsolaterally projecting lip of bone on lateral margin of premaxillary process of maxilla; dorsal surface of premaxillary process concave, bounded medially by crista transversalis; anterior inferior alveolar foramen of maxilla small; base of nasal process excavated anteriorly; anterior margin of nasal process vertical, overhangs premaxillary process; nasal process rugose, with small facet on anterior margin for articulation with nasal; teeth conical, tapering, with striated crowns and squarish cross-section posteriorly; Meckelian groove closed and extensively fused anteriorly and posteriorly; and a weak, discontinuous sulcus dentalis along most of the tooth row that is bounded medially by a sharp lip of bone.
Differs from Polychrus acutirostris, P. gutturosus and P. marmoratus in having conical, unicuspid teeth and greater overlap of the premaxilla onto the dorsal surface of the premaxillary process of the maxilla. Also differs from P. gutturosus and P. marmoratus in having a rugose nasal process of the maxilla. Also differs from P. acutirostris in having a more vertical nasal process and a less extended premaxillary process of the maxilla.
Etymology. charistikos, Gr. "giving freely," in reference to the permission given by the landowner, Jeff Oakland, for excavation of the specimens.
Description of holotype. The teeth of the maxilla are moderately robust, high-crowned and, as far as they are preserved, unicuspid, tapering apically. Weak labial and occasionally lingual striae are visible on the crowns despite heavy wear to the tips. The anterior teeth are slightly recurved, the more posterior teeth essentially straight (Figure 4.1–2). The palatal shelf is not wide. The palatine process is very weak, but there is a relatively large facet, developed on its dorsal surface and extending onto the main of the palatal shelf, that marks the articulation with the palatine. The superior alveolar foramen (SAF) is not well separated from the jugal groove, and it is set in a short gutter (see also below under Cypressaurus sp.). The nasal process, covered with irregular rugosities (interspersed with small foramina) above the labial foramina, has a nearly vertical anterior margin that even projects out over the premaxillary process (Figure 4.1). On the anterodorsal margin of the nasal process is a small but distinct facet that marks the articulation with the nasal. The maxillary anterior inferior alveolar foramen (AIAF) opens along this margin above the level of the premaxillary process.
Description of paratypes. Further information on the maxilla comes from PTRM 1841, 5373, and 5374. The crest for attachment of the vomer and the lamina transveralis anterior (Oelrich 1956)—here called the crista transversalis—is well developed, crossing the palatal shelf anteromedially, then extending nearly to the anterior end of the bone along its medial margin. The dorsal surface of the premaxillary process is concave, bounded by ridges medially and laterally (Figure 4.3). A second foramen—filled with sediment and continued by a small groove anteriorly—lies just lateral to the crista transversalis; it is probably for the subnarial artery (see Oelrich, 1956). A small lip of bone projects dorsolaterally to overhang part of the maxillary wall in all specimens. The premaxillary process is especially long, extending well beyond the anterior limit of the concavity on its dorsal surface (Figure 4.3). This anterior extension bears a facet dorsally for articulation with the premaxilla. Its anterior margin is somewhat concave in dorsal aspect. A ventral facet for articulation with the premaxilla is also present.
PTRM 5351 (Figure 4.4–5) is one of the largest dentaries. Though the bone itself is well preserved, only the bases of non-resorbed teeth remain; their crowns were likely broken during the screening process. Eighteen tooth spaces occupy 14.0 mm; the more anterior teeth are mesiodistally compressed (their bases are elliptical). The posterior teeth have broader bases and a squarish outline. There is a narrow, shallow, and discontinuous subdental gutter (sulcus dentalis) bordered medially by a sharp lip of bone. The dentary does not grow much taller posteriorly, and the dental parapet is low. Posteriorly, the dentary is broken at the level of the AIAF. Only the anterior portion of this foramen is preserved, and it is slit-like and narrow; a small groove extends anteriorly from it. The foramen is located in the upper half of the medial face of the dentary. The Meckelian groove is closed and extensively fused anteriorly. The anterior-most end of the dentary curves only slightly medially, not strongly and abruptly. There are five irregularly spaced labial foramina, the last of which is positioned between the twelfth and thirteenth teeth (from symphysis). There are a few fine, irregularly oriented grooves on the external surface of the dentary, which are probably artifactual. There is also a shallow, longitudinal groove on the ventral surface of the dentary. The ventral border of the dentary is nearly flat. The anterior opening of the Meckelian canal is highly restricted.
Other referred specimens supplement the description above. Tooth crowns in PTRM 5200 (only slightly worn) taper considerably toward their tips, even as posteriorly as tooth 13 (the last tooth preserved in this specimen). There is no indication at this point of tricuspid crowns. PTRM 1990 has two reasonably well-preserved teeth, at positions four and five (from symphysis). Each has a faint mesial crest descending the tooth crown, disappearing above the dental parapet. The fifth tooth has fine, vertical striations about midway up the lingual side of the crown and coarser striae on the labial side; the tip of the crown, though slightly worn, bears no indication of accessory cusps.
Remarks. The extensive (>50%) closure and fusion of the Meckelian groove in Polychrus charisticus is a derived feature it shares with the polychrotine taxa Anolis and Polychrus as well as with many iguanines and tropidurines (Etheridge and de Queiroz 1988). The extent of anterior fusion of the groove is matched, among living taxa, only by Anolis, Polychrus, and some Leiocephalus. The dentary of P. charisticus also appears (dorsoventrally) short posteriorly, but as the end of the bone is not preserved, it is not known how much taller it would become; nor is a dentary tooth count obtainable. Dentary tooth counts vary from 21 to 26 teeth in living Polychrus I have seen (26 in P. acutirostris, 24 in P. gutturosus, 21 in P. marmoratus).
The maxilla of Polychrus charisticus is distinctive and furnishes a number of useful characters. Especially noteworthy is the evident overlapping of the premaxilla onto the premaxillary process of the maxilla. In the few living iguanians where a dorsal overlap surface is evident on the premaxillary process—only Polychrus and the hoplocercine Enyalioides oshaughnessyi (where it is highly developed)—, the premaxilla has a posterolateral recess formed by the development above the primitive articulation surface of a flange that overlaps the maxilla. The premaxillary overlap was evidently more highly developed in P. charisticus than in P. marmoratus and P. gutturosus.
The moderately concave dorsal surface of the premaxillary process, and the dorsolaterally projecting lip of bone on its lateral margin, are seen in all examined Polychrus (see Appendix). A dorsal excavation is also found in many tropidurines (Plica, Uracentron, Tropidurus, Microlophus). In the latter taxa, however, the premaxillary process curves strongly medially toward its anterior end and lacks a dorsolaterally projecting ridge. A dorsal excavation is also seen in Enyalioides oshaughnessyi and many iguanines; but in iguanines the lateral crest bounding this dorsal excavation curves medially across the dorsal surface of the process. The excavation of the anterior base of the nasal process proceeds to such an extent in Polychrus marmoratus (Figure 4.6) and P. gutturosus (especially) that the lateral surface of the nasal process slightly overhangs the premaxillary process, as in P. charisticus (Figure 4.1). This was not observed in any other iguanian and is not well developed in P. acutirostris.
The superior alveolar foramen (SAF) is partially "roofed" in Polychrus charisticus, as in other disarticulated specimens of Polychrus, Anolis, many tropidurins (Microlophus, Plica, Tropidurus, Uracentron; less so in Uranoscodon), and phrynosomatines (see Appendix). In most other iguanians examined, including the agamid Uromastyx hardwickii and Leiocephalus, there is a gutter developed on the dorsal surface of the palatal shelf of the maxilla that extends far anteriorly toward the anterior end of the nasal process, and the superior alveolar nerve penetrates the maxilla in several places along the floor and anterior end of this gutter (see, e.g., Figure 11.3). Limited out-group comparison and the restricted distribution of the covered or "roofed" condition of the SAF in Iguanidae suggest that the former condition is derived and could serve to unite Anolis and Polychrus. The state of the SAF is unknown in leiosaurs and para-anoles (sensu Etheridge and de Queiroz 1988).
The irregular, nearly pustolose rugosities on the lateral surface of the nasal process in Polychrus charisticus also occur in some Anolis, some Polychrus (e.g., P. acutirostris), some Leiocephalus, some Phrynosoma, and some Laemanctus. In most iguanians, however, rugosities, when present, are restricted to the frontal and perhaps parietal. Extensive development of rugosities is considered derived in Iguania (Etheridge and de Queiroz 1988), but their complicated distribution in certain taxa (Anolis, Polychrus) disallows, at present, the use of this feature as an apomorphy uniting them.
The nasal facet on the anterior margin of the nasal process, seen in Polychrus charisticus, is also found in other Polychrus (Figure 4.6), corytophanines, Oplurus cuvieri, and Enyalioides oshaughnessyi. The derived nature of the maxilla in Anolis—which shows, for instance, a strong, low-angle canthal crest and a sharply medially bent nasal process—makes it difficult to ascertain whether this feature is primitive for Anolis, Polychrus, and Corytophaninae but lost due to transformation in Anolis. Stem representatives of Anolis would help to clarify this problem. In other iguanids, the nasal generally does not leave a distinct facet on the anterior margin of the nasal process; if it leaves a mark at all, it is an arcuate depression on the anteromedial surface of the nasal process. Out-group comparison with Agamidae and Sphenodon punctatus suggests that the presence of an anterior facet is primitive. It appears to have been lost, then, in Anolis, Pristidactylus torquatus (unknown in other leiosaurs), Chalarodon madagascariensis, Crotaphytinae, Phrynosomatinae, and Tropidurinae*.
Striations of the tooth crown are uncommon in Iguania, yet not as rare as commonly conceived. I have observed them labially and lingually in a number of Anolis (e.g., A. biporcatus, A. extremus, A. garmani), in all Polychrus, and in some Crotaphytus. They are also found in the early Eocene Parasauromalus olseni (see Gilmore 1928) and some Sceloporus (e.g., S. cyanogenys) and Tropidurus (e.g., T. hispidus). Distinct striations are interpreted as primitive for Polychrus and may independently have evolved in the other taxa listed above. The new species also likens Polychrus in the possession of a distinct if discontinuous sulcus dentalis that extends far posteriorly and whose medial border is a sharp lip of bone.
Several other features are primitive or of uncertain polarity and can do no more than exclude Polychrus charisticus from crown Anolis. For instance, the anterior margin of the nasal process of the maxilla is very steep in P. charisticus, as it is in many iguanids; but in Anolis, as well as some Polychrus (e.g., P. acutirostris, but not P. marmoratus or P. gutturosus), the margin rises more gradually posteriorly. As in Anolis and Polychrus, the crista transversalis is tall, a feature also seen in taxa like Phrynosomatinae.
In summary, Polychrus charisticus is united with Anolis and Polychrus by (1) extensive fusion of the Meckelian groove, both anteriorly and posteriorly, and (2) at least partial roofing of the SAF on the dorsal surface of the maxilla. Polychrus charisticus is united with living Polychrus by (1) extension of the premaxillary process of the maxilla, together with (2) dorsal overlap of the premaxillary process by the premaxilla, (3) slight overhang of the premaxillary process by the anterior margin of the nasal process, (4) the dorsolaterally projecting lip of bone on the lateral margin of the premaxillary process, and (5) lingual and labial tooth crown striations. The evident tuberculation of the dermal skull bones is not grossly inconsistent with this placement, as this feature appears in many taxa in both Anolis and Polychrus (though it cannot be concluded to be primitive for them). Known autapomorphies of P. charisticus are (1) the simplified tooth crowns, and (2) possibly the greater extent of overlap of the premaxillary process by the premaxilla. There are presently no features that would place the new species inside or outside crown Polychrus, and I have conservatively placed the new species in that taxon. If further information were to show the new species to lie outside crown Polychrus, charisticus could be removed.
Oddly, Polychrus is a predominantly South American taxon. Only one of six species, P. gutturosus, is found north of the Isthmus of Panama (according to the EMBL Database; see Köhler 2000). The sister-taxon of P. gutturosus, according to Frost et al. (2001), is P. femoralis, known exclusively from South America (EMBL Database; Lehr 2002). This small clade, in turn, was found to be the sister-taxon to remaining species of Polychrus. Polychrus gutturosus may be a fairly recent immigrant to Central America. If Polychrus charisticus is correctly referred, the story becomes more complex. Regardless of whether Polychrotinae* is paraphyletic (Frost et al. 2001), it seems that some members of the group had a longer history in North America than previously thought. P. charisticus would provide a good candidate for the estimation of divergence times in Iguania using molecular methods, provided that the position of Polychrus in Iguanidae is secured.
Size alone would not contravene the referral of Vertebra Type 1 (see below) to this species.
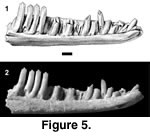 Referred specimens. PTRM 1822 (left dentary),
Figure 5; 2605 (right dentary fragment); 2743 (edentulous left dentary); 5231 (partial right dentary); 5334 (left dentary fragment); 5352 (partial right dentary); 5353 (right dentary fragment); 5354 (partial right dentary); 5724 (maxilla fragment)
Referred specimens. PTRM 1822 (left dentary),
Figure 5; 2605 (right dentary fragment); 2743 (edentulous left dentary); 5231 (partial right dentary); 5334 (left dentary fragment); 5352 (partial right dentary); 5353 (right dentary fragment); 5354 (partial right dentary); 5724 (maxilla fragment)
Description. The most complete specimen, PTRM 1822, is also one of the larger dentaries. There are spaces for 20 teeth in the ~10.8 mm of tooth row; at least one more tooth appears to have been present behind the twentieth. The teeth are relatively tall, and both anterior and posterior teeth are mesial-distally compressed (Figure 5). The eighth tooth (from symphysis) has distinct mesial and distal "shoulders," separated from the central cusp by very weak, vertical grooves that anticipate more posterior tooth morphology. Posterior teeth have strong mesial and distal accessory cusps, both separated from the central cusp by grooves. The crowns are parallel-sided (i.e., not flared). There is a slight concavity on the lingual side of each tooth, matched by a slight convexity on the labial side; thus, the cutting edges are directed somewhat lingually. The bases of the posterior teeth are slightly bulbous or protrusive. There is a weak sulcus dentalis anteriorly, but no subdental shelf or gutter posteriorly. The Meckelian groove is closed and fused, though it is impossible to determine precisely how far posteriorly, because the dentary is broken. A narrow, longitudinal groove extends anteriorly from the break, just ventral to the tooth row; this groove penetrates entirely through the medial wall of the dentary and may be artifactual. The external surface of the dentary is quite smooth; it is perforated by eight irregularly spaced labial foramina, the posterior-most of which is located at the level of the boundary between teeth 18 and 19 (from symphysis). The dentary does not curve strongly medially at its anterior end (i.e., it is not "hooked"). The anterior opening of the Meckelian canal is elongate.
Other specimens provide more information. In PTRM 5231, approximately the same size as PTRM 1822, the fifth and sixth teeth (from symphysis) have distinct "shoulders" separated by grooves. (The crown of the fourth tooth is broken; the third is resorbed.) PTRM 5353 is the medial segment of a small dentary, with tooth crown morphology similar to that of PTRM 1822. Interestingly, the crowns of the teeth are nearly translucent; this peculiarity is matched by PTRM 5354, a small partial dentary. PTRM 5354 also confirms the presence of posterior labial foramina. PTRM 2743, a highly weathered, edentulous partial dentary, preserved no indication of a medial, longitudinal groove on the dentary.
Remarks. This species is considered an iguanid because it possesses distinctly tricuspid teeth (grooves separating accessory cusps from the central cusp are present both labially and lingually), which are mesiodistally compressed and show pleurodont implantation. A number of other squamates have been described as having "tricuspid" teeth, such as scincoids, but in most of these taxa labial grooves bounding accessory cusps are not developed (the xantusiid Lepidophyma is an exception). In other taxa with distinctly tricuspid teeth, such as some teiids, they are mesiodistally expanded and tend to show considerable basal cementum. Most acrodont iguanians also have distinctly tricuspid teeth, but they show acrodont tooth implantation and are mesiodistally expanded. These remarks apply to other iguanids described below.
Iguanid MPH-1 is easily distinguished from Polychrus charisticus by several features. The teeth of the former are weakly tricuspid anteriorly (essentially unicuspid), and are closely spaced distally (widely spaced); the dental parapet is relatively higher (lower); the labial foramina extend far posteriorly (not far posteriorly); and the anterior opening of the Meckelian canal is elongate (constricted). Fusion of the Meckelian groove is a derived character in Iguania (Etheridge and de Queiroz 1988). Because this state was achieved independently multiple times, it is by itself of little systematic utility; as noted above, extensive fusion of the Meckelian groove appears also in Iguaninae, Anolis, Polychrus, and many tropidurines. There are otherwise few derived characters in the jaw elements that would constrain the affiliation of Iguanid MPH-1.
Size cannot disbar the referral of Vertebra Type 1 (see below) to this taxon.
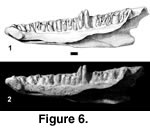 Referred specimens. PTRM 2602 (right dentary),
Figure 6
Referred specimens. PTRM 2602 (right dentary),
Figure 6
Description. The tooth row of this dentary is 15.0 mm long, and the depth of the dentary at the ultimate tooth is 4.6 mm. Twenty-three tooth positions, with one complete and one partial tooth in place, are preserved (Figure 6). The complete tooth is located in the latter half of the tooth row. It is relatively low-crowned. As preserved, the crown is relatively blunt, in part due to wear and to breakage of its mesial edge; there is no trace of a distal accessory cusp on the crown. Thus, cheek teeth in Iguanid MPH-2 were probably unicuspid or weakly tricuspid (this cannot be better constrained, due to the extent of wear). There is a narrow subdental shelf that spans much of the tooth row. The Meckelian groove is open, except for a space of ~2.5 mm in the anterior half of the dentary, where the dorsal and ventral flanges close on one another. A small groove (Figure 6) is present on the ventral surface of the dorsal flange at the level of the eighteenth tooth (from symphysis), marking the position of the AIAF, which would also then have been in contact with the splenial. The Meckelian groove is widely open posteriorly. The anterior end of the dentary is markedly inturned ("hooked").
Immediately posterior to the tooth row is a notch for the coronoid (Figure 6) which continues for a short distance onto the labial surface of the dentary. There was evidently no strong, anterior blade of the coronoid in this species, nor is there a large dorsal process of the dentary to brace the coronoid anteriorly. Several labial foramina penetrate the dentary's external surface; these tiny holes are irregularly spaced and extend >50% along the tooth row. The lateral surface of the dentary is otherwise smooth.
Remarks. The nearest relatives of Iguanid MPH-2 are obscure. Primitively, species of the phrynosomatine taxon Phrynosoma tend to have especially deep dentaries (Van Devender and Eshelman 1979), which may also apply to the Phrynosoma stem [e.g., Paraphrynosoma greeni (Holman 1987) from the early Oligocene of Colorado]. PTRM 2602 appears relatively deep, but the depth/length ratio is actually much closer to iguanids generally and does not suggest a relationship to Phrynosoma.
The small, deep, labial coronoid notch is likewise not currently diagnostic. A strong labial coronoid blade on the dentary is typical in members of Anolis (pers. obs.), Iguaninae (pers. obs.), Leiocephalus (Etheridge 1966; Pregill 1992; pers. obs.), and hoplocercines (Estes et al. 1988, figure 16; pers. obs.); a weaker one is found in some phrynosomatines and tropidurines (pers. obs.), and a blade is generally absent in members of Polychrus, Pristidacylus, Corytophaninae, Crotaphytinae, and Oplurinae that I have examined.
Finally, partial closure without fusion of the Meckelian groove is a derived feature also appearing in several iguanid clades. It is evident in a number of crotaphytines, Basiliscus, phrynosomatines, and oplurines (Etheridge and de Queiroz 1988) and in some liolaemins (Etheridge, personal commun., 2002). While on biogeographic grounds it is unlikely that Iguanid MPH-2 is a tropidurine or oplurine, this apomorphy cannot itself diagnose the species described here. Better specimens are also desirable before the species is formally named.
Type species. Tuberculacerta pearsoni sp. nov. (by monotypy)
Diagnosis. As for type and only known species.
Etymology. tuberculum, L., diminutive of tuber, L. "swelling, bulb," plus lacerta, L. "lizard," referring to the tuberculated snout of this squamate.
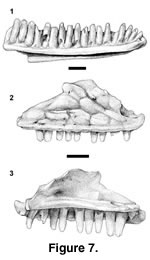 Holotype. PTRM 5296 (left dentary),
Figure 7.1
Holotype. PTRM 5296 (left dentary),
Figure 7.1
Paratypes. PTRM 1432 (left dentary fragment); 5326 (right dentary); 5355 (left dentary); 1429 (right maxilla), Figure 7.2–7.3
Locality and horizon. PTRM V89002, Chalky Buttes Member, Chadron Formation (late Eocene), North Dakota, USA
Distribution. Known only from type locality
Diagnosis. A small iguanid with parallel-sided to weakly tapering, tricuspid posterior teeth; >19 dentary teeth in total; subhorizontal intramandibular septum that extends far posteriorly; Meckelian groove narrow but open, faces ventrally near symphysis; lateral surface of maxilla adorned with large tubercles; sharp bend in the nasal process of the maxilla (see below); dorsal surface of the premaxillary process of the maxilla excavated; crista transversalis strong.
Etymology. Named in honor of Dean A. Pearson (PTRM), whose diligent work and kind permission has made this study possible.
Description of holotype. PTRM 5296 is a relatively complete left dentary, missing only its posterior-most end. The preserved tooth row is 8.0 mm long and contains 19 tooth spaces, with all but the first and sixth teeth in place. The teeth are moderately high crowned (Figure 7.1). The crowns are generally worn, but a couple of the posterior teeth retain mesial and distal cusps in addition to the main central cusp. The posterior crowns are parallel-sided to weakly tapering. Anteriorly there is a weak subdental shelf, but this shelf gradually disappears posteriorly. The Meckelian groove is entirely open; for most of its length it is directed ventromedially, but anteriorly it is directed ventrally. There is a prominent medial "hook" to the dentary at its anterior end. The intramandibular septum (IMS) extends far posteriorly and is present as far back as the dentary itself is preserved. The IMS is oriented nearly horizontally. Externally, there are six labial foramina, the last at the level of the fourteenth tooth.
Description of paratypes. PTRM 1432 and 5326 are smaller than PTRM 5296 and poorly preserved, but PTRM 5355, a still smaller specimen, provides more information on the pattern of tooth crown morphology in the dentary. The ninth and tenth teeth (from symphysis) are partially preserved, and weak grooves mesially and distally indicate that those teeth had at least weak "shoulders," if not distinct cusps. PTRM 5355 also has a somewhat better-developed subdental shelf posteriorly. All three of these specimens show a long, subhorizontal IMS, as seen in PTRM 5296.
PTRM 1429 is a well-preserved right maxilla (referred to the same taxon as the dentaries on the basis of size and tooth morphology). Twelve tooth positions and nine teeth are preserved (Figure 7.2–3). The teeth are unicuspid anteriorly, but the tenth and eleventh teeth are tricuspid, with parallel-sided to weakly tapering crowns; where precisely the transition occurs is difficult to estimate, because of wear on some more anterior teeth. The crowns are moderate in height. The nasal process is broad and shows a strong canthal crest (Figure 7.2); it is "bent," in other words, divided into an anterodorsally facing surface and a laterally facing one. Posteriorly, there is a facet on the medial surface of the nasal process for articulation with the prefrontal. Anteriorly, the nasal process is continuous with the well-developed crista transversalis. The nasal process shows large, well-developed dermal rugosities, which presumably indicate the margins of epidermal scales, down to just above the main row of labial foramina, of which there are six. The premaxillary process is broken medially at the level of the subnarial arterial foramen. The dorsal surface of the process is excavated. The AIAF lies just dorsal and medial to this pit, at the base of the nasal process. The palatine process, as preserved, is insignificant, but the sharp-bordered facet visible on its dorsal surface (Figure 7.3) and the tapering of the palatal shelf medial to this facet suggest that it has been broken. Its original extent is unknown. The SAF is "roofed" (see above under Polychrus charisticus) and opens posteriorly near the end of the nasal process (Figure 7.3). The posterior end of the maxilla is broken, but it retains part of the moderately deep groove for articulation with the jugal. This groove extends anteriorly to the level of the SAF.
Remarks. The elongate IMS of Tuberculacerta pearsoni is noteworthy. To evaluate this feature in iguanids, I measured two lengths in each of over 50 isolated dentaries, including representatives of all major clades except Liolaemini. The first length is that from the symphysis to the end of the IMS; the second, from the symphysis to the end of the tooth row. The ratio between these measurements was ≥0.69 in all phrynosomatines (usually >0.80) and >0.80 in Anolis, Pristidactylus torquatus, Oplurinae, and Leiocephalus. A ratio of <0.69 was found in all other clades, including Polychrus, Corytophaninae, Crotaphytinae, Enyalioides oshaughnessyi, Iguaninae, and Tropidurini (Uracentron, Uranoscodon, Microlophus, Tropidurus, Plica). In cases where two specimens of the same species were examined (Phrynosoma asio, Gambelia wislizenii), the ratio differed by no more than 7%. In the four agamids examined, the ratio was between 0.60 and 0.70. An extended IMS—defined here as one showing a ratio of greater than ~0.7—is tentatively interpreted as independently synapomorphic of some part of Polychrotinae*, Oplurinae, Leiocephalus, and Phrynosomatinae, pending evaluation in Liolaemini. The posterior end of the most complete dentary in the sample, PTRM 5296, is not preserved; one can nevertheless say that the IMS extends to the level of the nineteenth dentary tooth, further than in all iguanids with a "short" IMS (ratio less than ~0.7).
In the maxilla of Tuberculacerta pearsoni, as in all phrynosomatines examined, the SAF is "roofed." As discussed above, this morphology appears to be synapomorphic of Anolis and Polychrus, of parts of Tropidurinae*, and of Phrynosomatinae.
The nasal process of the maxilla of Tuberculacerta pearsoni is distinctive in being strongly bent medially and continuous with the crista transversalis at its anterior base. A bend like this, which corresponds to a strong canthal crest, is well developed in all examined members of Phrynosomatinae with the exception of some Phrynosoma (P. cornutum, P. platyrhinos). As discussed below, the nasal process in these species of Phrynosoma departs strongly from the primitive condition. Among other iguanians examined, a sharp bend is present in Tropidurini but is weak in Leiocephalus and oplurines; it is weak or absent in many Polychrus (P. gutturosus, P. marmoratus), Crotaphytinae, Iguaninae, and Enyalioides oshaughnessyi. In the case where a sharp bend is absent, the nasal process of the maxilla is essentially a vertical flange that may curve slightly medially at its dorsal terminus. On the other hand, a sharp bend is present in Anolis (even better developed here) and in Corytophaninae; in Anolis, the anterodorsal surface of the nasal process is continuous with the crista transversalis, as in Phrynosomatinae, whereas in Corytophaninae the bend occurs high and in a nearly coronal plane, such that the (small) dorsally facing surface is not continuous with the crista. Outgroup comparison suggests that the presence of a sharp bend is derived in Iguanidae where it occurs. Thus, the distribution within Iguania of a sharp bend in the maxilla suggests that it is primitive for Phrynosomatinae (and also Tropidurini and Anolis) and has been transformed beyond recognition in some members of the eponymous clade Phrynosoma. Absence in T. pearsoni of a closed and fused Meckelian groove (primitive for Tropidurinae and Polychrotinae*) suggests in turn that it is related to Phrynosomatinae; this conclusion is consistent with the extended IMS and "roofed" SAF discussed above.
Rugosities on the nasal process of the maxilla, as seen in Tuberculacerta pearsoni, also occur in some species of Phrynosoma. Rugosities on the dermal bone in Phrynosoma generally extend far beyond the frontal, regardless of whether they reach the nasal process, which is highly reduced in many species. Among other iguanids examined, some members of Polychrotinae* and Crotaphytus have the best-developed dermal rugosities. Pregill (1992) has also discussed them in some species of Leiocephalus but determined them not to be primitive for that clade; Etheridge and de Queiroz (1988) and Lang (1989) furthermore noted them in Laemanctus serratus. Extreme development of rugosities on the dermal skull bones may be a synapomorphy of Phrynosoma or part of its stem, and their presence in T. pearsoni might suggest that this species belongs on that stem.
This evidence tying Tuberculacerta pearsoni to the stem of Phrynosoma is tenuous. In any case, the new taxon lacks at least two synapomorphies of (crown) Phrynosoma, namely, a medially facing anterior extent of the Meckelian groove (Van Devender and Eshelman 1979) and a high dentary depth/length ratio. Many species of Phrynosoma furthermore possess an open Meckelian groove (P. modestum is an exception), a characteristic that distinguishes them from other phrynosomatines. An open Meckelian groove is primitive for Iguania (Etheridge and de Queiroz 1988), but it is important to determine whether its occurrence in Phrynosoma is symplesiomorphic or secondary. If closure of the Meckelian groove was acquired twice in Phrynosomatinae—once in the Uta-Urosaurus-Sceloporus clade and once in "sand lizards" (sensu Frost and Etheridge 1989)—then an open Meckelian groove in T. pearsoni only excludes it from these clades; if closure is a synapomorphy of Phrynosomatinae and reversed in Phrynosoma, then the open Meckelian groove could unite T. pearsoni with Phrynosoma. The (uncertain) phylogenetic position of Petrosaurus is crucial to addressing this question.
A maxilla of Tuberculacerta pearsoni preserving the palatine process is desirable. If it is a phrynosomatine, as the evidence above suggests, it should have a strong and triangular palatine process. In most phrynosomatines (excepting Petrosaurus mearnsi and some Sceloporus like S. clarkii and S. serrifer) the palatine process is moderate to large in size and triangular. This morphology also obtains in oplurines, crotaphytines, and many tropidurines (although in Leiocephalus and Uranoscodon the process tends to be blunt), but contrasts with the morphology in Polychrotinae*, Corytophaninae, Crotaphytinae, Iguaninae, and Enyalioides oshaughnessyi, where the process is blunt, rounded, and poorly developed. One can conclude, minimally, that a strong, triangular palatine process is primitive for Phrynosomatinae.
In summary, Tuberculacerta pearsoni possesses a larger suite of characters uniting it with Phrynosomatinae than with any other taxon, including (1) an elongate IMS, (2) a sharp bend in the nasal process of the maxilla, creating an anterodorsally facing surface that is continuous with the crista transversalis, and (3) a "roofed" SAF. Within Phrynosomatinae, T. pearsoni is tenuously united with Phrynosoma by the well-developed rugosities of the facial bones. The open Meckelian groove does not contravene this placement.
Tuberculacerta pearsoni has at least one autapomorphy that distinguishes it from all extant phrynosomatines examined, namely, the dorsal excavation of the premaxillary process of the maxilla. A slight depression is evident in the premaxillary process of Cophosaurus texana and Urosaurus graciosus, but it is not as well developed as in T. pearsoni; in most phrynosomatines examined, the premaxillary process is flat and may or may not slope slightly ventrolaterally.
Thus far, the earliest record of Phrynosomatinae seems to be the early Oligocene Paraphrynosoma greeni (Holman 1987), which was also suggested to lie on the stem of Phrynosoma.
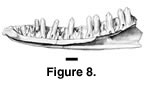 Referred specimens. PTRM 5198 (right dentary),
Figure 8; 2054 (left dentary fragment); 2594 (partial left dentary); 5202 (partial right dentary); 2095 (left maxilla fragment); 5356 (right maxilla fragment); 5401 (left maxilla fragment); ?5396 (left maxilla fragment)
Referred specimens. PTRM 5198 (right dentary),
Figure 8; 2054 (left dentary fragment); 2594 (partial left dentary); 5202 (partial right dentary); 2095 (left maxilla fragment); 5356 (right maxilla fragment); 5401 (left maxilla fragment); ?5396 (left maxilla fragment)
Description. PTRM 5198 is a right dentary missing only its posterior-most end (Figure 8). It preserves 23 tooth spaces (with nine teeth); the length of the tooth row as preserved is 10.9 mm. The teeth are moderate in height. There are well-developed mesial and distal accessory cusps on the crowns of the posterior teeth, but the crowns are not flared. These cusps are much weaker on the more anterior teeth but nevertheless visible on tooth nine. The posterior crowns do not abut one another. The posterior-most tooth is somewhat shorter than the anterior-most, reflecting a decrease in tooth size toward the back of the tooth row. Five middle teeth occupy a length of 5.4 mm. Anteriorly, a moderately developed subdental gutter is present, which is reduced, but still present, posteriorly. The IMS extends to under the twentieth tooth space (9.2 mm along the tooth row). The Meckelian groove is open along the entire dentary; however, it narrows considerably in the middle. The anterior end of the dentary turns abruptly medially (it is "hooked"). On the external surface, there is a linear indentation, running longitudinally just below the parapet of the jaw. There are five labial foramina, the last located at the level of the fourteenth tooth space. The remaining specimens provide little more information on dentary morphology, although it is worth noting that PTRM 2594, nearly the same size as PTRM 5198, lacks an external, longitudinal indentation.
The maxilla of cf. Aciprion sp. is not represented by equally well-preserved material. (Maxillae were referred to the same taxon as the dentaries on the basis of size and tooth morphology.) PTRM 2095 has essentially the same tooth morphology as the dentaries described above. It is a middle fragment of maxilla; it is quite worn and does not preserve the nasal process. The roof of bone covering the posterior course of the superior alveolar foramen is broken, giving the foramen the superficial appearance of being set in a "gutter"; it was not. PTRM 5356 is a slightly more posterior fragment that preserves the anterior part of the jugal groove, which is only moderately deep.
Remarks. Cf. Aciprion sp. compares closely in many details with the equi-sized Aciprion formosum from the early Oligocene of Colorado (YPM 4663, YPM-PU 10015). Cf. Aciprion sp. had >23 teeth in the dentary, versus 25 in YPM-PU 10015. Tooth morphology, especially the position and nature of the transition from unicuspid to tricuspid crowns, and tooth spacing are similar in the two taxa. The only significant dental difference between the two appears to be tooth crown height, which is lower in cf. Aciprion sp. The two taxa additionally share a similar symphyseal structure. A. formosum does differ from cf. Aciprion sp. in that it has only a weak subdental shelf; additionally, in A. formosum the Meckelian groove is closed (but not fused) for several millimeters in the anterior half of the dentary, while in cf. Aciprion sp., the groove merely narrows.
As in Aciprion formosum (see Estes 1983), there are few features of cf. Aciprion sp. that permit ready phylogenetic placement. The apparently high IMS/tooth row length ratio (~0.8?) is suggestive of affinities with Phrynosomatinae/Oplurinae/Tropidurini or with Anolis (see above under Tuberculacerta pearsoni).
 Referred specimens. PTRM 5357 (partial left dentary),
Figure 9; 5178 (partial left dentary); 5267 (left dentary fragment)
Referred specimens. PTRM 5357 (partial left dentary),
Figure 9; 5178 (partial left dentary); 5267 (left dentary fragment)
Description. PTRM 5357 is the posterior segment of a left dentary (Figure 9). It is generally well worn, with bone edges and tooth crowns rounded. Ten tooth positions and seven teeth are preserved. The teeth are moderately high-crowned. The anterior-most tooth was either unicuspid or weakly tricuspid. The teeth become more strongly tricuspid posteriorly; the crowns begin to flare and actually contact one another to form a continuous cutting surface. Due to the incomplete preservation, it is unclear where along the dentary the transition in tooth crown morphology occurs. Five middle teeth occupy 4.6 mm. There is a very weak subdental shelf anteriorly. The Meckelian groove is open throughout the preserved length of dentary. PTRM 5178 and 5267 provide no further details.
Remarks. Iguanid MPH-3 is very similar to cf. Aciprion sp. By comparison with the latter, however, Iguanid MPH-3 is distinguished by having more closely spaced teeth and slightly flared tooth crowns that, in the posterior part of the jaw, contact one another. The IMS appears to be posteriorly extensive in this taxon, but better-preserved and more complete specimens must be found before any systematic determination can be made.
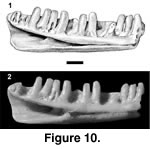 Referred specimen. PTRM 2041 (partial left dentary),
Figure 10
Referred specimen. PTRM 2041 (partial left dentary),
Figure 10
Description. PTRM 2041, the middle segment of a left dentary, is heavily worn. Fourteen tooth positions and nine teeth are preserved. The teeth are worn to rounded stumps, so no distinct morphology is apparent (Figure 10); they are, however, not closely spaced. There is the slightest of subdental gutters along most of the length of the tooth row. The Meckelian groove gently closes anteriorly. The IMS extends posteriorly to under the tenth preserved tooth.
Remarks. Iguanid MPH-4 is poorly preserved, and little can be said about the characteristics of this morphotype. As the tooth crowns are worn away, tricuspid teeth cannot be used to refer it to Iguanidae; instead, complete closure (without fusion) of the Meckelian groove must be used (I am unaware of any non-iguanid that shows this feature). It is distinguished from most other taxa described here by gentle closure, without fusion, of the Meckelian groove and from Cypressaurus sp. (see below) by the more robust teeth.
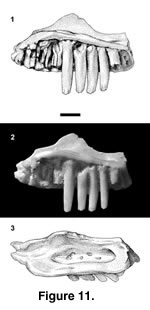 Referred specimens. PTRM 2623 (partial left maxilla),
Figure 11; 5321 (dentary fragment)
Referred specimens. PTRM 2623 (partial left maxilla),
Figure 11; 5321 (dentary fragment)
Description. PTRM 2623 is the medial portion of a left maxilla. The four complete teeth are high-crowned, with ~50% of their height projecting above the parapet of the jaw. The tooth crowns are slightly worn, but they are distinctly tricuspid. The crowns taper slightly toward their apices (Figure 11.1–2). The palatine process of the maxillary shelf is weak. There is a small groove along the dorsal surface of its anterior margin that represents the articulation with the palatine. The SAF is set in a gutter (Figure 11.3; see also discussion under Polychrus charisticus and Tuberculacerta pearsoni). The jugal groove is quite deep; it extends anteriorly to the posterior end of the gutter but is separated from it by a small ridge. The external surface of the nasal process is smooth and has four labial foramina as preserved.
The dentary fragment has three teeth with the same general tooth form as PTRM 2623; the crowns are somewhat worn. The ventral portion of the dentary is not preserved.
Remarks. The monotypic Cypressaurus hypsodontus Holman (1972) was described on the basis of two partial dentaries from the medial Chadronian of Saskatchewan. PTRM 2623 is referred to Cypressaurus on the basis of its high tooth crowns, a derived feature. In C. hypsodontus, the central cusp of each tooth appears higher than in PTRM 2623, but this difference could be the result of wear. Also in C. hypsodontus, the tooth crowns do not appear to taper toward the apex (see Holman 1972, figure 2), unlike in PTRM 2623. More and better specimens are desirable to clarify whether these differences indicate the presence of two different taxa.
![]()