 New handsome fungus beetles (Coleoptera: Coccinelloidea: Anamorphidae, Endomychidae) from European amber of the Upper Eocene
New handsome fungus beetles (Coleoptera: Coccinelloidea: Anamorphidae, Endomychidae) from European amber of the Upper Eocene
Article number: 21.1.6A
https://doi.org/10.26879/832
Copyright Paleontological Society, February 2018
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 5 November 2017. Acceptance: 7 February 2018
{flike id=2141}
ABSTRACT
Five new genera and eight new species of handsome fungus beetles of the superfamily Coccinelloidea are described and imaged from Baltic and Bitterfeld amber: Gramboale prutenorum gen. et sp. nov., Kleinzaches germanicianus gen. et sp. nov., Palaecoryphus viktori gen. et sp. nov., Giltine ampeensis gen. et sp. nov., Symbiotes borussiaeorientalis sp. nov. (Anamorphidae), Laima andreei gen. et sp. nov. (Endomychidae: Endomychinae), Trochoideus koenigsbergicus sp. nov., and T. resinatissimus sp. nov. (Endomychidae: Pleganophorinae). Additional specimens of Glesirhanis bercioi Shockley and Alekseev and Phymaphoroides antennatus Motschulsky (Endomychidae: Leiestinae), from Baltic amber are also reported. A key to the known handsome fungus beetles genera with formally described species from Eocene ambers is provided.
Vitalii I. Alekseev. Department of Zootechny, Kaliningrad State Technical University, Sovetsky av. 1, 236000 Kaliningrad, Russia. alekseew0802@yahoo.com
Wioletta Tomaszewska*. Museum and Institute of Zoology, Polish Academy of Sciences; Wilcza 64, 00-679 Warszawa, Poland. wiolkat@miiz.waw.pl
Keywords: fossil resin; inclusions; new species; new genera; Paleogene; Eocene
Final citation: Alekseev, Vitalii I. and Tomaszewska, Wioletta. 2018. New handsome fungus beetles (Coleoptera: Coccinelloidea: Anamorphidae, Endomychidae) from European amber of the Upper Eocene. Palaeontologia Electronica 21.1.6A 1-23. https://doi.org/10.26879/832
palaeo-electronica.org/content/2018/2141-new-coccinelloidea-from-eocene
Copyright: February 2018 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/152941A8-052F-4134-9E98-ABC4CAC8F01B
INTRODUCTION
The fossils of handsome fungus beetles in a broad sense (Tomaszewska 2000, 2005, 2010; Shockley et al., 2009a) are poorly known and studied, and only three extinct endomychid species have been described from European Eocene amber so far. The fossil record of Endomychidae sensu lato has been summarized in Shockley and Alekseev (2014). After recent changes in cucujiform beetle classification (Robertson et al., 2015), resulting among others in separation of Endomychidae sensu stricto, and raising Anamorphidae, Mycetaeidae, and Eupsilobiidae as independent families, the study of fossil members of these groups is of special importance and of interest.
In the current paper, new fossil taxa of handsome fungus beetles from the extant genera Trochoideus Westwood, 1833 (Endomychidae: Pleganophorinae) and Symbiotes Redtenbacher, 1849 (Anamorphidae), and also from five extinct genera of both families are described, illustrated, and discussed. As a summary of morphology and for practical purposes, a key to the known handsome fungus beetles genera with formally described species from Eocene ambers is here provided (Appendix 1).
The genus Trochoideus is distributed in all the tropics of the world and is represented by 20 described species: 19 extant and 1 fossil (Shockley et al., 2009a). Nine species are known in South and Central America (T. americanus Buquet, 1840, T. goudoti Guérin-Méneville, 1857, T. boliviensis Strohecker, 1978, T. coeloantennatus Strohecker, 1943, T. globulicornis Joly and Bordon, 1996, T. masoni Strohecker, 1978, T. peruvianus Kirsch, 1876, T. venezuelensis Joly and Bordon, 1996, T. mexicanus Strohecker, 1978); two inhabit Madagascar (T. dalmani Westwood, 1838, T. wasmanni Strohecker, 1959); one is described from Africa (T. sansibaricus Kolbe, 1897); six species occur in South-Eastern Asia (T. bicolor Csiki, 1909, T. microphthalmus Wasmann, 1922, T. minutus Csiki, 1909, T. mirabilis Strohecker, 1958, T. fea Gorham, 1896, T. tonkineus Strohecker, 1980); and one species possesses an almost cosmopolitan distribution (T. desjardinsi Guérin-Méneville, 1838); while one known fossil species has been described from the Madagascan copal (T. cruciatus [Dalman, 1825]).
The genus Symbiotes includes five described species (Shockley et al., 2009a). Two species are endemic to the Palaearctic region (S. armatus Reitter, 1881, and S. latus Redtenbacher, 1849), and two are endemic to the Nearctic region (S. duryi Blatchley, 1910 and S. impressus Dury, 1912). Symbiotes gibberosus (Lucas, 1846) is native to the Palaearctic, but has now become widespread across the Nearctic region, presumably due to human activity (Shockley and Tomaszewska, 2007).
MATERIAL AND METHODS
Nine specimens from the private collection of Christel and Hans Werner Hoffeins (Hamburg, Germany), and one specimen from the private collection of the first author (Kaliningrad, Russia) were examined during this study. The amber samples currently deposited in the private collection of Christel and Hans Werner Hoffeins (CCHH) will be given to the Senckenberg Deutsches Entomologisches Institut in Müncheberg, Germany (SDEI) as part of the institute’s amber collection. The type from the private collection of first author (CVIA) will be deposited in the Paleontological Institute, Russian Academy of Science (Moscow) for permanent preservation. The origin of the amber pieces is presented in Table 1.
The bioinclusions from Baltic amber were obtained from commercial sources in Poland and in the Kaliningrad region, Russia. The specimens from Bitterfeld amber were collected in the Goitzsche coal mine, Sachsen-Anhalt, Germany. All CCHH pieces (except for No 53-2) were cut, polished, and embedded in blocks of transparent polyester resin according to the procedure of Hoffeins (2001). The specimen from the CVIA collection (AWI-133) and No 53-2 from the CCHH are without any further fixation.
The photos of specimen from piece 1219-3 [CCHH] were taken with Nikon Coolpix 4500 camera mounted on a Wild M3Z stereomicroscope. All other photos were taken with a Zeiss AxioCamICc 3 digital camera mounted on a Zeiss Stemi 2000 stereomicroscope. Reconstructions were made based on freehand drawing during examination of the original specimens by the first author. The figures were edited using Adobe Photoshop CS8.
The following sources were used for the generic attribution and comparison with extant and extinct taxa: Gerstaecker (1858), Walton (1912), Strohecker (1953, 1958), Joly and Bordon (1996), Sasaji (1990), Tomaszewska (2000, 2004), and Shockley (2006, 2010).
SYSTEMATIC PALAEONTOLOGY
Based on Tomaszewska (2000), all beetle specimens from the amber pieces studied during this project are referred to the family Endomychidae sensu lato, as they have the following set of morphological characters: head with fronto-clypeal suture; antennae with club composed of three antennomeres; procoxae separated by prosternal process; elytral epipleura well developed; abdomen with five ventrites; ventrite 1 as long as two following ventrites combined, and postcoxal lines on abdominal ventrite 1 absent; tarsi 4- or 3-segmented.
Family ANAMORPHIDAE Strohecker, 1953
Remarks. Beetle specimens from amber pieces No. 1799-1, No. 181-1, No. 1222-3, No. AWI-133, and No. 1799-2 possess all characters of family Anamorphidae, such as: mesocoxal cavities widely closed outwardly by meso- and metaventrite; head without antennal grooves; antenna composed of 10 or 11 antennomeres, with terminal antennomere simple not modified; second labial palpomere subcylindrical; tarsal formula 4-4-4, 3-3-3, or sometimes 3-3-4, with simple tarsomeres and sometimes appendiculate tarsal claws; and body almost always distinctly pubescent (Tomaszewska, 2000; Robertson et al., 2015).
Genus GRAMBOALE gen. nov.
zoobank.org/CFB5BF8C-442B-4397-9315-8CAEBAFDBB04
Type species. Gramboale prutenorum sp. nov.
Diagnosis. Gramboale can be differentiated from all other extant and fossil anamorphids by the following combination of characters: body comparatively large (length more than 3 mm), ovoid and convex, and dorsal surface without vestiture; procoxal cavities open externally; antennal club simple, symmetrical; pronotum finely bordered laterally, lacking lateral sulci and lateral carinae, and with posterior margin weakly bisinuate; elytra punctate-striate without explanate margins; tarsal claw with sharp internal tooth. Moreover, the new genus differs from fossil Discolomopsis Shockley, 2006 by not having explanate margins of elytra; an absence of dorsal vestiture; and by having 4-segmented tarsi. Gramboale can be distinguished from extant Rhymbomicrus Casey, 1916 by posterior pronotal margin not lobed medially; from Anamorphus LeConte, 1878, Micropsephus Gorham, 1887-99, and Bystus Guérin-Méneville, 1857 by an absence of lateral sulci on pronotum; and from Symbiotes Redtenbacher, 1849, Clemmus Hampe, 1850, and Exysma Gorham, 1887-1899 by an absence of pronotal lateral carinae. The new genus clearly differs from Bystodes Strohecker, 1953, Mychothenus Strohecker, 1953, and Idiophyes Blackburn, 1895 by having glabrous elytra, and pronotum without lateral sulci; from Anagaricophilus Arrow, 1922, Cyrtomychus Kolbe, 1910, and Geoendomychus Lea, 1922 by having 11 segmented antennae, and absence of lateral pronotal sulci; from Dexialia Sasaji, 1970, and Dialexia Gorham, 1887-1899 by elytra with regular rows of punctures, and 11 segmented antennae. Among Anamorphidae Gramboale is most similar to Australian genus Erotendomychus Lea, 1922 in having pronotum bordered laterally and without sulci, mesoventrite with intercoxal process rounded and prominent anteriorly, tarsi 4-segmented, and 3-segmented antennal club; but it differs from Erotendomychus in short-oval body form, by elytral punctures forming regular or almost regular rows, by five abdominal ventrites, and absence of dorsal pubescence.
Etymology. The name of the new genus is derived from “gramboale” –“beetle” in the Old Prussian language. The gender is masculine.
Composition. The new genus is monotypic.
Gramboale prutenorum sp. nov.
Figure 1
zoobank.org/9222D44D-A06C-4458-A4F0-D3F31BBCE428
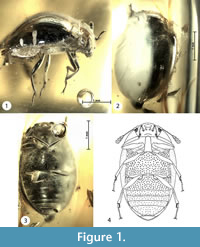 Type material. Holotype: No. 1799-1 [CCHH]. Presumably male. The beetle inclusion is preserved in a polished piece of transparent amber with a yellowish shade. The small amber piece is embedded in a block of polyester resin (total measurements are: 14 x 7 x 6 mm). Syninclusions absent. Left half of the beetle is obscured by milky foam in the surrounding amber.
Type material. Holotype: No. 1799-1 [CCHH]. Presumably male. The beetle inclusion is preserved in a polished piece of transparent amber with a yellowish shade. The small amber piece is embedded in a block of polyester resin (total measurements are: 14 x 7 x 6 mm). Syninclusions absent. Left half of the beetle is obscured by milky foam in the surrounding amber.
Type strata. Baltic amber. Eocene.
Type locality. Russia, Kaliningrad region, the Sambian (Samland) peninsula, Yantarny settlement (formerly Palmnicken).
Etymology. The species name is derived from the Latin name of the West Baltic tribe Pruteni or Prutheni (Prussians). The epithet is a noun in plural form and in genitive case.
Diagnosis. As stated for new genus.
Description. Body length = 3.4 mm; width (at widest point in the middle part of elytra) = 1.8 mm; height = 1.38 mm. Rounded, convex habitus; dorsal surface shiny and glabrous; body including appendages uniformly black.
Head wide, retracted into prothorax up to surface of compound eyes, punctate, finely shagreened; puncture diameters are 0.5 times that of one compound eye facet; width of interspaces 1.0-3.0 times puncture diameter. Punctation on frons slightly sparser than on clypeus. Fronto-clypeal suture present. Eyes rounded, slightly prominent. Antennae relatively long, extending to base of elytra, composed of 11 antennomeres with conspicuous, pubescent club composed of three antennomeres; antennomere 1 obovate. Terminal maxillary palpomere as long as antennomere 2, obconical. Terminal labial palpomere pointed, rounded-triangular; penultimate palpomere subcylindrical.
Pronotum convex, transverse, 0.47 times as long as wide; with rounded posterior and anterior angles; finely bordered laterally. Posterior margin of pronotum weakly bisinuate, anterior margin weakly concave. Pronotum finely shagreened anteriorly, coarsely and moderately densely punctate, with width of interspaces 1.5-2.0 times puncture diameter laterally and with width of interspaces 3.0-4.0 times puncture diameter medially. Prosternal process wide. Procoxal cavities open externally, closed internally. Mesoventrite with intercoxal process rounded and prominent anteriorly between mesocoxae.
Elytra rounded, convex, shiny, glabrous, with weak humeri, punctate-striate. Punctures (rounded on elytral disc and oval laterally), forming regular or almost regular rows or shallow furrows. Sutural striae indistinct. Scutellum visible, weakly transverse, rounded posteriorly. Epipleura well developed, almost reaching apex of elytra. Hind wings not apparent.
Meso- and metacoxae widely separated (by distance equal almost two times and three times longitudinal coxal diameter, respectively). Mesocoxal cavities closed laterally. Abdomen with five ventrites; ventrite 1 as long as two following ventrites combined. Ventrites 2, 3, and 4 subequal in length. Apex of terminal ventrite widely rounded. Postcoxal lines on ventrite 1 absent. Thoracic punctation dense and large, abdominal punctation (especially on ventrites 2-5) sparser and distinctly smaller.
Femur subclavate; meso- and metafemur with shallow groove on inner margin. All tibiae simple (without angulations or projections), weakly curved, covered with short pubescence. Tarsal formula 4-4-4. Tarsi long, with simple cylindrical segments, pubescent (tarsomere 1 bearing tuft of very dense and long setae ventrally). Tarsomere 4 as long as tarsomeres 1-3 combined. Tarsal claws long (0.3 x as long as tarsomere 4), curved, symmetrical, toothed basally.
Remarks. The holotype is most likely a male, based on the presence of modifications on the legs in the form of tufts of setae on tarsomere 1. Genitalia are internal and obscured in this specimen, and are generally impossible to differentiate in specimens embedded in amber.
Genus KLEINZACHES gen. nov.
zoobank.org/2FF026B9-3060-476F-AD50-EB194F12ABF0
Type species. Kleinzaches germanicianus sp. nov.
Diagnosis. Combination of small overall body size (1.45 mm), antennae composed of 10 antennomeres, long lateral sulci reaching three-quarters of pronotal length, dorsum convex and covered with long semierect pubescence, pronotum bordered laterally and with weakly bisinuate posterior margin, regularly punctured elytra, long ventrite 1, tarsal formula 3-3-4 and appendiculate tarsal claws distinguishes Kleinzaches from all currently described extant and extinct genera of Anamorphidae.
Etymology. The name of the new genus is derived from “klein Zaches” –a character in the fairy-tale “Klein Zaches, genannt Zinnober” (“Little Zaches, acclaimed as Zinnober”). The tale was written by Ernst Theodor Amadeus Hoffmann (1776-1822), a famous Prussian author of fantasy and Gothic horror, born in Königsberg (now Kaliningrad). The gender of the new genus is masculine.
Composition. The new genus is monotypic.
Kleinzaches germanicianus sp. nov.
Figure 2
zoobank.org/4707AE6B-3FF6-4E2C-9973-DC535DA4273B
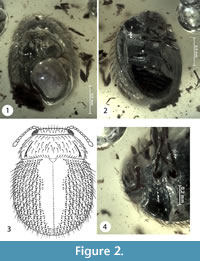 Type material. Holotype: No. 181-1 [CCHH]. Sex unknown. The beetle inclusion is preserved in a polished piece of transparent amber, yellowish in color. The small amber piece is embedded in a block of polyester resin (total measurements are 9 x 8 x 3 mm). Syninclusions consist of numerous small wood pieces and three gas bubbles. The right hind leg and terminal antennomere of right antenna of the beetle are missing.
Type material. Holotype: No. 181-1 [CCHH]. Sex unknown. The beetle inclusion is preserved in a polished piece of transparent amber, yellowish in color. The small amber piece is embedded in a block of polyester resin (total measurements are 9 x 8 x 3 mm). Syninclusions consist of numerous small wood pieces and three gas bubbles. The right hind leg and terminal antennomere of right antenna of the beetle are missing.
Type strata. Bitterfeld amber. Eocene.
Type locality. Germany, Sachsen-Anhalt, Goitzsche (Bitterfeld-Wolfen).
Etymology. The specific epithet refers to the region of the specimen’s origin and deposition: “germanicianus” in Latin literally means “stationed in Germany.”
Diagnosis. As stated for new genus.
Description. Body length = 1.45 mm; width (at widest point in middle of elytra) = 1.25 mm. Elytral length (along suture) = 1.05 mm. Habitus: broadly oval, dorsally convex; dorsal surface shiny, covered with long erect hairs; body uniformly dark brown with appendages and ventrites 2-5 lighter.
Head retracted into prothorax up to position of compound eyes, pubescent. Compound eyes rounded, weakly prominent. Fronto-clypeal suture distinct. Antennae composed of 10 antennomeres, extending to half length of pronotum, with loose club composed of three antennomeres. Antennomere 8 oblong, almost twice as long as wide; antennomere 9 as long as wide; antennomere 10 symmetrical, oblong and pointed. Terminal labial and maxillary palpomeres oblong, weakly pointed.
Pronotum transverse (2.5 times as wide as long), widest within one-third of its length; bordered laterally and anteriorly. Lateral sulci long, narrow, spanning posteriormost three-fourths of pronotal length. Pronotum covered with sparse, irregular, semirecumbent pubescence. Pronotal posterior margin almost straight. Procoxal cavities externally open, internally closed. Prosternal process somewhat triangular with rounded angles, distinctly bordered throughout. Mesocoxal cavities widely closed laterally.
Elytra rounded, convex, with base slightly wider than posterior margin of pronotum, covered with long pubescence, with small strial punctures, arranged in almost regular rows. Distance between punctures in each row 1.0-2.0 times larger than diameter of each point, and intersticial width is about three times diameter. Spaces between rows with regular long erect setae, separated from each other by distance shorter than length of each seta. Elytral epipleura long, reaching ventrite 5; with rows of small punctures at outer margin.
Abdomen with five ventrites; apex of terminal ventrite widely rounded; postcoxal lines absent. Ventrite length proportions (measured along midline) according to formula: 17: 6: 6: 6: 11. Metacoxae widely separated by intercoxal process of ventrite 1; process is twice as wide as longitudinal metacoxal diameter and straight anteriorly. Distance between irregular and fine punctures on abdominal ventrites is 1.0-3.0 times larger than diameter of each puncture.
Legs with femora thickened apically. Tibiae slender. Tarsi long, with simple cylindrical segments. Tarsal formula 3-3-4. Metatarsomeres 1-3 almost equal in length; metatarsomere 4 longest, almost as long as tarsomeres 1-3 combined. Tarsal claws quite long (0.3 times as long as apical tarsomere), symmetrical, appendiculate.
Remarks. The periscutellar area (scutellum, central part of pronotal base and basal part of elytra) is invisible because of gas bubbles.
Genus PALAECORYPHUS gen. nov.
zoobank.org/868C55C1-D8BA-4EE7-81E5-5D929EE325BE
Type species. Palaecoryphus viktori sp. nov.
Diagnosis. This new genus may be separated from all currently described extant and extinct genera of Anamorphidae by the following combination of characters: small overall body size (1.6 mm); dorsal surfaces of body covered with long semierect pubescence; pronotum comparatively short with truncate posterior margin, with basal sulcus deep and lateral sulci long; elytra irregularly punctate; ventrite 1 long; tarsi 4-segmented; and antennae composed of 11 antennomeres.
Moreover, Palaecoryphus differs from fossil Discolomopsis Shockley, 2006 in having very narrow (not widely explanate) lateral margins of elytra and pronotum, and tarsal formula 4-4-4. Palaecoryphus is similar to extant Asymbius Gorham, 1896, Coryphus Csiki, 1902, Clemmus Hampe, 1850, and Exysma Gorham, 1887-99 in having rather straight (not lobed medially) posterior margin of pronotum, and ovoid habitus, but clearly differs from these genera in all tarsi being 4-segmented. Palaecoryphus additionally differs from Asymbius in having scutellar striae that are not widely curved in front; and from Coryphus by its subquadrate penultimate antennomere. New genus resembles also Catapotia Thomson, 1860 in having 11 antennomeres and tarsal formula of 4-4-4, but differs in presence of lateral sulci and truncate posterior margin of pronotum. Asiatic genera Mychothenus Strohecker, 1953, Bystodes Strohecker, 1953, Idiophyes and Dexialia Sasaji, 1970 can be distinguished from Palaecoryphus in having 10 antennomeres and different form of pronotum.
Etymology. The new genus-group name Palaecoryphus is composed of “palaeo” (from Greek ‘palaios’ meaning old, ancient) and “Coryphus”, the name of the extant, habitually similar anamorphid genus from Papua New Guinea. The gender is masculine.
Composition. The new genus is monotypic.
Palaecoryphus viktori sp. nov.
Figure 3.1-3
zoobank.org/D7C9518E-9FEB-4275-A3C6-E0233F0A80BE
 Type material. Holotype: No. 1222-3 [CCHH]. Sex unknown. The beetle inclusion is preserved in a polished piece of transparent amber with a yellowish shade. The small amber piece is embedded in a block of polyester resin (total measurements are 16 x 10 x 5 mm). Syninclusions consist of one mite. Underside and right side of the beetle are obscured by milky foam within the surrounding amber.
Type material. Holotype: No. 1222-3 [CCHH]. Sex unknown. The beetle inclusion is preserved in a polished piece of transparent amber with a yellowish shade. The small amber piece is embedded in a block of polyester resin (total measurements are 16 x 10 x 5 mm). Syninclusions consist of one mite. Underside and right side of the beetle are obscured by milky foam within the surrounding amber.
Type strata. Baltic amber. Eocene.
Type locality. Russia, Kaliningrad region, the Sambian [Samland] peninsula, Yantarny settlement [formerly Palmnicken].
Etymology. Patronymic, this new species is dedicated to the son of the first author, Viktor V. Alekseev.
Diagnosis. As stated for new genus.
Description. Body length 1.6 mm; width (at widest point within anterior one-third of elytra) 1.1 mm. Broadly oval, convex habitus; dorsal surface covered with long pubescence; body and head uniformly dark, almost black; antennae and most part of legs brown, with palpi and tarsi paler.
Head retracted into prothorax up to point of compound eyes, which are rounded, weakly prominent. Antennae relatively long, extending to base of elytra; composed of 11 antennomeres, and pubescent; basal antennomeres slender, followed by loose club, composed of three antennomeres. Antennomere length proportions follow formula: 6: 5: 3.3: 3.3: 3.3: 3.3: 3.3: 3.3: 5: 5: 9. Antennomere 9 subquadrate, antennomere 10 transverse, antennomere 11 obconical. Terminal maxillary palpomere spindle-shaped and pointed; terminal labial palpomere cylindrical, pointed.
Pronotum transverse (0.3 times as long as wide), widest at middle; basal transverse sulcus deep; lateral sulci long (reaching two-thirds of pronotal length) and deep. Anterior and posterior angles rounded, anterior angles weakly produced. Posterior margin of pronotum truncate, anterior almost straight, lateral sides arcuate. Pronotum coarsely and densely punctate with interspaces 0.5-1.0 times puncture diameter. Pronotum covered with long, semierect setae (as long as 3.0-4.0 times puncture diameter).
Elytra rounded (1.1 times longer than wide; 4.25 times longer than pronotum) and convex; irregularly coarsely and densely punctate, and pubescent like pronotum; with weak humeri; wider than posterior margin of pronotum; with narrowly flattened margins. Sutural striae distinct from elytral apex to near scutellum, becoming evanescent in scutellar area; not curved near scutellum. Scutellum visible, strongly transverse (2.5 times wider than long). Epipleura present, well developed, reaching posterior margin of ventrite 1. Hind wings not apparent.
Procoxae narrowly separated, meso- and especially metacoxae widely separated. Mesocoxal cavities closed laterally. Abdomen with five ventrites; ventrite 1 as long as four following ventrites combined. Ventrites 2, 3, 4 and 5 subequal in length. Apex of terminal ventrite rounded. Postcoxal lines absent.
Legs with femora clavate, tibiae simple (without any angulations or projections), widened toward apex, covered with short pubescence. Tarsal formula 4-4-4. Tarsi long, with simple cylindrical segments. Tarsomere 4 as long as tarsomeres 2-3 combined. Tarsal claws 0.3 times as long as tarsomere 4, simple, symmetrical.
Remarks. Antero-dorsal part of the head is not visible, so the presence of fronto-clypeal suture in this specimen cannot be confirmed.
Genus GILTINE gen. nov.
zoobank.org/7627E41D-B10B-4DBF-B7C9-FD1F41E7E852
Type species. Giltine ampeensis sp. nov.
Diagnosis. Giltine can be separated from all currently described extant and extinct genera of Anamorphidae by the following combination of characters: small overall body size (1.7 mm); convex and pubescent dorsum; pronotum bordered laterally and biconcave at base, with basal pronotal sulcus shallow and weakly marked, and with long lateral carinae; elytra punctate-striate; 4-segmented tarsi with simple tarsal claws.
Etymology. This new genus-group name is a theonym and derived from Giltinė - Lithuanian goddess of death, sister of Laima. The gender is feminine.
Composition. The new genus is monotypic.
Giltine ampeensis sp. nov.
Figure 3.4-6
zoobank.org/8B52DA23-278F-4C1F-9C7C-B7AF32C8E552
Type material. Holotype: No. AWI-133 [CVIA]. Sex unknown. The complete beetle inclusion is preserved in a polished, small, triangular piece of transparent amber, yellowish in color (total measurements are 9 x 7 x 4 mm). Syninclusions include four stellate hairs. The piece was obtained from a commercial source in the Yantarny settlement (Kaliningrad region) in August 2015. It was polished by hand, thus allowing dorsal and lateral views of the included beetle.
Type strata. Baltic amber. Eocene.
Type locality. Russia, Kaliningrad region, the Sambian (Samland) peninsula, Yantarny settlement (formerly Palmnicken).
Etymology. The species name “ampeensis” is derived from “Ampe”–the old name of Palmnicken (now Yantarny settlement).
Diagnosis. As stated for new genus.
Description. Body length 1.7 mm; width (at widest point in anterior one-third of elytra) 1.25 mm. Broadly oval, dorsally convex habitus; dorsal surface shiny, with long, curved setae; body uniformly dark, almost black, with tarsi slightly lighter.
Head retracted into prothorax to point of compound eyes, pubescent. Eyes rounded, prominent. Antennae relatively long, extending to base of elytra; with loose club composed of three antennomeres. First antennomere of antennal club elongate; second antennomere of antennal club as long as wide; terminal antennomere symmetrical, oblong, and pointed.
Pronotum transverse (2.5 times as wide as long), widest within posteriormost one-third of length; bordered laterally and anteriorly. Lateral carinae long, arcuate, spanning posterior two-thirds of pronotal length. Pronotum covered with sparse and large setiferous punctures. Distance between punctures 3-5 times as large as diameter of each puncture. Setae long, curved, semirecumbent. Pronotal posterior margin biconcave.
Elytra rounded, convex, wider than base of pronotum, with long pubescence, with margins explanate and strial punctures arranged in almost regular rows. Distance between punctures in row 1.5-2.0 times as large as diameter of each puncture. Spaces between rows (interstices) with regular setiferous punctures (slightly smaller than punctures in rows). Setae on elytra semirecumbent, long, arranged in rows on interstices, as long as distance between setae. Scutellum visible, rounded, impunctate, transverse (1.5 times wider than long).
Legs with femora flattened. Tibiae slender. Tarsi long, with simple cylindrical segments. Tarsal formula 4-4-4. Metatarsomeres 1 and 4 longest, 1.5 times longer than metatarsomeres 2 or 3. Tarsal claws 0.3 times as long as tarsomere 4, simple, symmetrical.
Remarks. The specimen is obscured ventrally by milky foam within the surrounding amber, and it is impossible to determine the number of ventrites, the length of epipleura, the details of ventral side of thorax, and the number and form of antennomeres of antennal stalk; the antenna consists of 10 or 11 antennomeres. Genitalia are internal and obscured in this specimen, and are generally impossible to differentiate in specimens embedded in amber.
Genus SYMBIOTES Redtenbacher, 1849
Symbiotes borussiaeorientalis sp. nov.
Figure 4.1-3
zoobank.org/DB91C913-B619-447D-930A-E816A976F371
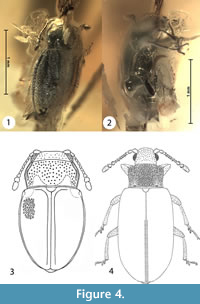 Type material. Holotype No. 1799-2 [CCHH], female (terminal parts of genitalia are visible; apices of elytra without small tubercles and other modifications). The beetle inclusion is preserved in a polished piece of transparent amber with an orange shade and the amber piece is embedded in polyester resin (total measurements are 11 x 9 x 4 mm). Syninclusions absent.
Type material. Holotype No. 1799-2 [CCHH], female (terminal parts of genitalia are visible; apices of elytra without small tubercles and other modifications). The beetle inclusion is preserved in a polished piece of transparent amber with an orange shade and the amber piece is embedded in polyester resin (total measurements are 11 x 9 x 4 mm). Syninclusions absent.
Type strata. Baltic amber. Eocene.
Type locality. Russia, Kaliningrad region, the Sambian (Samland) peninsula, Yantarny settlement (formerly Palmnicken).
Etymology. The specific epithet refers to the region of the specimen’s origin and is formed after the Medieval Latin “Borussia orientalis” (Eastern Prussia).
Remarks. The newly described species from Baltic amber was assigned to the genus Symbiotes based of following combination of morphological characters: body form subparallel; antenna composed of 11 antennomeres; procoxal cavities open externally; pronotum with posterior margin truncate and lateral margins raised; pronotal carinae not reaching posterior margin; and tarsi 4-segmented.
Diagnosis. Symbiotes borussiaeorientalis differs from all known congeners in having coarsely and irregularly punctured elytra with distinct subsutural striae only, and scarce, poorly visible dorsal pubescence. Newly described species is 1.6 mm long, thereby smaller than most extant representatives of Symbiotes (1.7-2.4 mm), except S. gibberosus, which is 1.5-1.8 mm long. Additionally, new fossil species can be distinguished from S. duryi and S. gibberosus by having smooth (not denticulate) lateral pronotal margins; from S. latus by its less transverse pronotum (2.5 times wider than long in S. latus), wider elytra, and dark color; from S. gibberosus by less convex body, and absence of pits at base of lateral sulci; from S. duryi, S. armatus, and S. latus by presence of subsutural stria without distinct punctation; and from S. armatus by widely rounded scutellum.
Description. Body uniformly dark brown, elongate oval, moderately convex, shiny, scarcely pubescent. Total body length 1.6 mm, width (at widest point) 0.75 mm.
Head deeply retracted in pronotum, transverse; with fine and very sparse punctation. Eyes large, prominent. Fronto-clypeal suture distinct, straight. Antennae comparatively long, extending to base of elytra; composed of 11 antennomeres, with loose club composed of three antennomeres. Antennomere length proportions follow formula: 2.5: 2.0: 1.0: 1.0: 1.0: 1.0: 1.0: 1.0: 1.5: 2.5: 3.5. Antennomeres 3-8 subequal in length and form, short and cylindrical; antennomere 9 triangular; antennomere 10 as long as wide; terminal antennomere subcylindrical, rounded apically, almost 2.0 times as long as wide. Terminal maxillary palpomere elongate, subcylindrical, tapering.
Pronotum widest at middle, strongly transverse (2.0 times wider than long); pronotal anterior and posterior margins almost straight, anterior angles obtuse, and basal angles right-angled. Lateral margins of pronotum arcuate, broad, raised, and lateral edges smooth. Median region convex, base with well-marked transverse sulcus, lateral sulci large, triangularly longitudinal, reaching anteriorly nearly one-half length of pronotum. Pronotal disc punctate: coarsely and moderately densely at sides (interspaces 1.0-2.0 times puncture diameter), more finely and sparsely near middle (interspaces 2.0-4.0 times puncture diameter).
Elytra comparatively long (3.0 times longer than pronotum), oval, decidedly wider than prothorax, attaining their greatest width within about one-third of their basal length; with explanate margins. Elytral disc coarsely and densely punctate; elytral striae absent except subsutural ones. Punctures irregular, interspaces 0.2-0.5 of diameter, denser laterally and sparser medially. Elytral epipleura pubescent. Subsutural stria distinct, broadly curved near scutellum and reaching middle of elytral base. Elytral apices conjointly rounded. Scutellum visible, transverse (about two times wider than long), widely rounded posteriorly.
Legs with procoxal cavities open externally. Metacoxae transverse, widely separated. Femur widest in middle, more than twice as wide as tibia. All tibiae simple. Tarsal formula 4-4-4. Tarsomeres simple, cylindrical; tarsomere 4 longest. Tarsal claws simple, almost 0.3 times as long as tarsomere 4.
Abdomen with five ventrites. Ventrite 1 as long as 2 and 3 combined; ventrites 2, 3, and 4 subequal in length; ventrite 5, 1.5 times longer than 4. Apex of terminal ventrite widely rounded.
Family ENDOMYCHIDAE Leach, 1815
Subfamily ENDOMYCHINAE Leach, 1815
Remarks. The beetle specimen from amber piece No. 1222-5 possesses characters of the subfamily Endomychinae (according to Robertson et al., 2015, this subfamily includes now Endomychus Panzer, 1795 and all genera previously classified in the subfamily Stenotarsinae), such as: body densely pubescent (glabrous in most species of Endomychus); pronotum with broad lateral, raised margins (except for Saula Gerstaecker, 1858 and Endomychus); head without occipital file and antennal grooves; anterior margin of pronotum without stridulatory membrane; tarsi pseudotrimerous with lobed tarsomere 1 and 2; and mesocoxal cavities open laterally (Tomaszewska, 2000; Robertson et al., 2015). Therefore we assign it to the subfamily Endomychinae.
Genus LAIMA gen. nov.
zoobank.org/C3B47AF1-BDF5-440C-9E96-082312AB7CA7
Type species. Laima andreei sp. nov.
Diagnosis. Laima can be diagnosed from all known extant Endomychinae genera by the following combination of characters: black elongate body; irregularly punctate and densely pubescent elytra; pronotum much narrower than base of elytra, with anterior angles not produced, and lateral sulci with two small tubercles; and sutural striae distinct only apically.
Laima closely resembles extant Saula (elongate body, irregularly punctate and uniformly colored elytra, dense pubescence, pronotum narrower than elytral base with anterior angles not produced); but Laima differs from Saula in having black body, smaller size, smooth (not crenulate) lateral margins of pronotum, rounded scutellum and five abdominal ventrites. Moreover, Laima can be distinguished from Danae Reiche, 1847 by pronotal form (without broad, raised lateral margins, without produced anterior angles, and with lateral sides not arcuate), and by presence of five abdominal ventrites; from Stenotarsus Perty, 1832 by elongate body with subparallel elytra, and not protruding anterior angles of pronotum; from Tragoscelis Strohecker, 1953 by having not as stout antennae and lacking deflexed acute anterior angles of pronotum; from Ectomychus Gorham, 1887 by having symmetrical antennal club, deep basal sulcus on pronotum, and lacking protruding pronotal angles. Laima can be easily distinguished from only other fossil endomychid beetle known from Baltic amber, with similar length and elongate body form, Phymaphoroides antennatus Motschulsky, 1856, based mainly on shape of ninth antennomere (simple in Laima, and strongly enlarged in Phymaphoroides).
Etymology. This new genus-group name is a theonym and derived from Laima, the name of a goddess from the Baltic pagan religion (the goddess of fortune, destiny, fate, pregnancy and childbirth). The gender is feminine.
Composition. The new genus is monotypic.
Laima andreei sp. nov.
Figure 4.4, Figure 5.1-3
zoobank.org/06EC1ADB-DB38-4ABF-9A26-6146FA80AC69
 Type material. Holotype: No. 1222-5 [CCHH]. Sex unknown. The beetle inclusion is preserved in a polished piece of transparent amber with a yellowish shade. The small amber piece is embedded in a block of polyester resin (total measurements are 12 x 8 x 5 mm). Syninclusions consist of numerous small wood pieces and filaments.
Type material. Holotype: No. 1222-5 [CCHH]. Sex unknown. The beetle inclusion is preserved in a polished piece of transparent amber with a yellowish shade. The small amber piece is embedded in a block of polyester resin (total measurements are 12 x 8 x 5 mm). Syninclusions consist of numerous small wood pieces and filaments.
Type strata. Baltic amber. Eocene.
Type locality. Russia, Kaliningrad region, the Sambian (Samland) peninsula, Yantarny settlement (formerly Palmnicken).
Etymology. The specific epithet is a patronym for Dr. Karl Erich Andrée, a German geologist and paleontologist, Director at the Geological-Paleontological Institute, and in charge of the amber collections of Albertina (Königsberg University).
Diagnosis. As stated for new genus.
Description. Body length 3.0 mm; width (at widest point in middle part of elytra) 1.25 mm; elongate; dorsal surface irregularly punctate and densely pubescent, uniformly black.
Head transverse, fine and comparatively scarcely punctate (with interspaces 1.0-2.5 times puncture diameter). Fronto-clypeal suture distinct. Eyes rounded, prominent, coarsely facetted. Antennae relatively long, pubescent, extending to base of elytra; composed of 11 antennomeres with loose club composed of three antennomeres. Antennomere length proportions follow formula: 10: 6: 5: 3: 3: 3: 5: 5: 7: 7: 11. Terminal maxillary palpomere cylindrical, longitudinal, apically rounded; penultimate palpomere short, transverse, broader than terminal one. Terminal labial palpomere longer than wide, pointed apically.
Pronotum transverse, 0.66 times as long as wide; weakly convex; with widely rounded anterior and rectangular posterior angles. Anterior pronotal margin slightly convex, posterior margin almost straight; posterior part of pronotum with deep transverse basal sulcus; lateral sulci shallow, each with two small tubercles basally. Pronotal disc coarsely and moderately densely punctate with interspaces 0.3-0.5 times puncture diameter. Prosternal process not visible due to position of legs. Procoxae are very close to each other; mesocoxae separated by distance 0.5 times of mesocoxal diameter; metacoxae broadly separated, with distance equal to coxal diameter. Hypomeron smooth.
Elytra elongate (1.7 times longer than wide; 3.6 times longer than pronotum), subparallel; with weak humeri; densely pubescent and irregularly punctate (like pronotum). Sutural striae distinct apically and reaching from apex to basal one-third of elytra. Scutellum visible, transverse (twice as long as wide), widely rounded posteriorly. Hind wings present.
Legs with tibiae simple and femora cylindrical. Tarsal formula 4-4-4. Tarsi pseudotrimerous; first and second tarsomeres lobed; tarsomere 4 as long as all previous segments combined; tarsomere 1 twice as long as tarsomere 2; tarsomere 2 and 3 almost equal in length. Tarsal claws simple, curved, almost one-quarter length of tarsomere 4.
Abdomen with five ventrites; ventrite 1 as long as three following ventrites combined, without postcoxal lines. Ventrites 2, 3, and 4 subequal in length. Apex of terminal ventrite widely rounded.
Subfamily PLEGANOPHORINAE Jacqueline du Val, 1858
Remarks. Beetle specimens from amber pieces No. 1506-1 and No. 53-2 possess characters of the endomychid subfamily Pleganophorinae, such as: body usually pubescent; tarsi 4-segmented, simple; antenna composed of 4-5 antennomeres with the terminal antennomere modified/enlarged (Tomaszewska 2000, Robertson et al. 2015). Therefore, we assign them to this subfamily.
Genus TROCHOIDEUS Westwood, 1833
Trochoideus koenigsbergicus sp. nov.
Figure 5.4-5
zoobank.org/EFF25E20-6F35-471D-83E8-613C18C79550
Type material. Holotype No. 1506-1 [CCHH], sex unknown. The beetle inclusion is preserved in a polished piece of transparent amber with a yellowish shade and the amber piece is embedded in polyester resin (total measurements are 20 x 15 x 6 mm). The syninclusions consist of three fagacean stellate hairs, one Symphypleona (Collembola), and one Nematocera (Diptera) that is 1.2 mm in length.
Type strata. Baltic amber. Eocene.
Type locality. Russia, Kaliningrad region, the Sambian (Samland) peninsula, Yantarny settlement (formerly Palmnicken).
Etymology. The species name is derived from Königsberg (now Kaliningrad, Russia), the German name of the capital of the former East Prussia.
Remarks. The new species from Baltic amber is diagnosed as belonging to genus Trochoideus based on following morphological characters: body weakly elongate; broad head with protuberant eyes; pronotum convex, widest near middle of its length and then narrowed posteriorly, with posterior angles right-angled; and elytra elongate-oval with weak, rounded humeral angles.
Diagnosis. Trochoideus koenigsbergicus differs from all extant and extinct species of Trochoideus in having antennae composed of five antennomeres with antennomeres 2-4 equal in proportions. At 3.1 mm long, it is smaller than most members of Trochoideus. Additionally, it can be distinguished from T. mirabilis Strohecker, 1958, and T. desjardinsi Guérin-Méneville, 1838 by having pronotum widest in anterior one-third; from T. coeloantennatus Strohecker, 1943, and T. globulicornis Joly and Bordon, 1996 by presence of distinct parasutural elytral striae; from T. mexicanus Strohecker, 1978, and T. dalmani Westwood, 1838 by an absence of central longitudinal sulcus and long lateral sulci on pronotum; from T. americanus Buquet, 1840, T. goudoti Guérin-Méneville, 1857, and T. peruvianus Kirsch, 1876 by disk of pronotum being rugosely punctate at sides; and from T. dalmani Westwood, 1838, T. wasmanni Strohecker, 1959, and T. cruciatus (Dalman, 1825) by having dark, monochromatic elytra.
Description. Body elongate, convex, uniformly dark brown, with short pubescence all over body and legs. Total body length 3.1 mm, width (at widest point) 1.5 mm.
Head with coarse and dense punctation (interspaces as large as puncture diameter). Eyes large, prominent. Fronto-clypeal suture straight. Clypeus covered with dense, relatively long, semierect hairs; and clypeal shape flat, transverse, with subparallel sides. Antennae short, extending to mid-length of pronotum; composed of five antennomeres, claviform. Antennomere length proportions follow formula: 2.0: 1.0: 1.0: 1.0: 5.7. Terminal antennomere rounded, weakly widened apically, 2.8 times as long as wide, with few dispersed, long, erect hairs (additional to background pubescence).
Prothorax transverse, 1.3 times wider than long; pronotal anterior margin concave with raised forward, widely-rounded anterior angles; posterior angles right-angled. Lateral margins of pronotum arcuate from anterior margin to anterior one-third, and slightly converging posteriorly within posterior two-thirds. Posterior margin of pronotum weakly bisinuate. Pronotum convex, bordered laterally, without central longitudinal sulcus; lateral sulci distinct, shallow, triangular, as long as one-quarter of pronotal length, connected by deep transverse basal sulcus. Pronotal disc coarsely and moderately densely punctate medially (interspaces 1.0-2.0 times puncture diameter), rugosely punctate at sides.
Elytra elongate (1.3 times as long as wide, widest at end of anterior one-third), convex; 2.4 times longer than prothorax. Elytral disc finely, densely punctate, with laterally visible pubescence. Punctures irregular, slightly longitudinally oval, interspaces widths equal to 0.5-2.0 puncture diameters, punctures denser basally and sparser apically. Parasutural striae distict, fine, entire. Elytral apices conjointly rounded. Scutellum visible, transverse (about 2 times wider than long), widely rounded posteriorly.
Legs with tarsal formula 4-4-4. Tarsomeres 1-3 with comb of long hairs ventrally; tarsomere 1 and 4 longest. Tarsal claws simple, long, almost as long as 0.5 times length of tarsomere 4.
Remarks. The characters of the ventral part of the specimen (distance between pro-, meso-, and metacoxae, coxal cavities, number of ventrites, and structure of protibia) are unfortunately not visible on the specimen because the dense milky cover in the surrounding amber.
Note. The extant Trochoideus species have sexually dimorphic antennal structure. The antennae of males and females can differ in form, proportions and number of antennomeres (e.g., four in males and seven in females). The single studied specimen of new species cannot be sexually determined, but sexual dimorphism in the fossil Trochoideus species is very probable.
Trochoideus resinatissimus sp. nov.
Figure 5.6-7, Figure 6.1-2
zoobank.org/2456F93D-89AA-4850-8452-99A33CF5E6B4
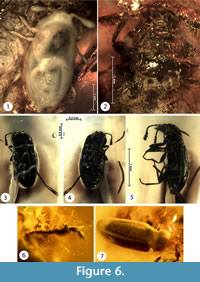 Type material. Holotype No. 53-2 [CCHH], sex unknown. The beetle is included in a polished piece of transparent amber with an orange shade. The amber was not subjected to any additional fixation. Measurements of the amber piece are 20 mm x 17 mm x 5 mm. The amber piece was possibly subjected to thermal and high-pressure processing in an autoclave.
Type material. Holotype No. 53-2 [CCHH], sex unknown. The beetle is included in a polished piece of transparent amber with an orange shade. The amber was not subjected to any additional fixation. Measurements of the amber piece are 20 mm x 17 mm x 5 mm. The amber piece was possibly subjected to thermal and high-pressure processing in an autoclave.
Type strata. Bitterfeld amber. Eocene.
Type locality. Germany, Sachsen-Anhalt, Goitzsche (Bitterfeld-Wolfen).
Etymology. The epithet of the new species is Latin adjective “resinatus” (resinous) in the superlative.
Diagnosis. Trochoideus resinatissimus differs from congeneric and probably coeval species described above from Baltic amber, based on antennal structure (strongly transverse antennomere 4 and egg-shaped antennomere 5) and larger body.
Description. Body elongate, convex, uniformly dark. Total body length 4.1 mm, elytral length 2.7 mm, width (at widest point in middle of elytra) 2.0 mm.
Compound eyes prominent and coarsely facetted. Antennae short, extending to middle of pronotum; composed of five antennomeres, claviform. Antennomere length proportions follow formula: 3.2: 1.2: 1.4: 1.0: 8.0. Penultimate antennomere strongly transverse (three times as wide as long). Terminal antennomere egg-shaped, rounded apically, 2.0 times as long as wide. Terminal labial palpomere almost spherical. Maxillary palp with terminal palpomere as long as penultimate one, oblong and acuminate. Anterior angles of pronotum widely-rounded. Metathorax coarsely and densely punctate. Distance between metacoxae wider than distance between mesocoxae. Elytra laterally with fine, irregular, slightly longitudinally oval punctures. Apex of elytra with distinct and fine parasutural stria. Legs with tarsal formula 4-4-4. Abdomen with six ventrites, and ventrite 1 longest.
Remarks. The specimen is dorsally covered with dense milky cover, so the characters of upper surface are not visible (except lateral and apical parts of the elytra).
Note. According to the note given under the description of Trochoideus koenigsbergicus (above) from Baltic amber, the new species from Bitterfeld amber could theoretically be the male of the same, widely distributed Eocene species. The thickened antennomere 4 of T. resinatissimus and different form of other antennomeres could be interpreted as sexual characters only. However, the question about reliable specific differences between specimens from Baltic and Bitterfeld amber would require further study of additional fossil material from both deposits.
Subfamily LEIESTINAE Thomson, 1863
Glesirhanis bercioi Shockley and Alekseev, 2014
Figure 6.3-5
Material examined. One well-visible, complete specimen with the collection number 1219-2 [CCHH], Baltic amber, Yantarny, Kaliningrad Region, Russia. Body length of the beetle 2.1 mm. The amber piece is embedded in a block of polyester resin (total measurements are 16 x 6 x 4 mm). Syninclusions include two tarsi of an insect (possible Diptera, Nematocera) and two parts of legs of an Arthropoda.
Note. The second known specimen of the species is smaller than holotype, which is 2.5 mm long.
Phymaphoroides antennatus Motschulsky, 1856
Figure 6.6-7
Material examined. One complete specimen with the collection number 1219-3 [CCHH], Baltic amber, Yantarny, Kaliningrad Region, Russia. Body length of the beetle 3.04 mm. The amber piece is embedded in a block of polyester resin (total measurements are 12 x 10 x 6 mm). Syninclusions absent. The amber piece was subjected to thermal and high-pressure processing in an autoclave. Left and ventral parts of specimen are obscured by a dense milky cover in the surrounding amber.
Note. The important characters are unfortunately not visible or insufficiently visible on the specimen. The re-description and exact subfamilial placement of Phymaphoroides are still impossible based on this specimen. The discernible enlarged antennomere 9 with deep concavity on its inner edge looks like that of the subfamily Danascelinae Tomaszewska, 2000. In the future, detailed comparison of a better visible Phymaphoroides specimen with the newly described Laima gen. nov. is recommended, due to observed similarities between both studied specimens.
However, without a study of the type specimen of Phymaphoroides antennatus, the proper subfamilal placement of this genus cannot be resolved. And unfortunately the location of the type material described by Motschulsky from Baltic amber in the Menge collection [West-Preußisches Provinzial-Museum Danzig], among which is the holotype of Ph. antennatus, is unknown, precluding a reinvestigation of this fossil. The original description lacks many characters important for comparison (Shockley and Alekseev, 2014) making subsequent conclusions about Phymaphoroides speculative.
DISCUSSION
Despite fossil Endomychidae sensu lato being included in several major catalogues (Klebs, 1910; Spahr, 1981; Carpenter, 1992; Strohecker, 1953; Shockley et al., 2009a), there are only seven species that have been properly named and fully described until now. Among these only three species have been described from European ambers (Shockley and Alekseev, 2014). This situation proves that despite recent interest in insect fossils (especially those preserved in resins), the knowledge of fossil faunas of handsome fungus beetles is very poor. This gap in our knowledge impedes making conclusions about the origin of both families (Anamorphidae and Endomychidae) and the superfamily Coccinelloidea itself.
Research to date has suggested the origin of the family Endomychidae sensu lato to have occurred in the Eocene (Carpenter, 1992), or during the Paleocene (65-54.8 Ma) (Kirejtshuk, 2000; Kirejtshuk and Nel, 2009). However, estimates by Ponomarenko (2002), indicated the origin during the Late Cretaceous, and the report of Endomychidae in Burmese (98 Ma) and Lebanese amber (125 Ma) by Poinar and Poinar (2008) further support a much older origin for this group. The recent discovery of the Endomychidae sensu stricto members in Burmese amber, confirms at least the Cretaceous origin of Coccinelloidea (Tomaszewska and Ślipiński, in preparation).
The growing number and diversity of handsome fungus beetle inclusions reported in Baltic amber alone indicates that past diversity may have been highly underestimated and needs extensive research. This work may also shed some light on conditions in the ancient forest that produced the amber deposits.
Relatively little is known about anamorphid and endomychid natural history so far (Shockley et al., 2009b). Two types of habitats are preferred by most species: 1) the subcortical region of standing or fallen dead trees, where they are closely associated with the fruiting bodies of macrofungi; and 2) leaf litter, where they feed on the spores and hyphae of microfungi. Most handsome fungus beetles are mycophagous, feeding on hyphae, spores and spore-bearing tissue, and the supporting tissues of fungal fruiting bodies. Within Endomychidae sensu stricto numerous species exhibit a special form of mycophagy known as sporophagy (feeding on spores), but obligate sporophagy has been hypothesized for all species of the family Anamorphidae (including Symbiotes), at least in the larval stages. Several species of Symbiotes sometimes feed on oyster mushrooms, Pleurotus ostreatus. The representatives of this genus are collected from tree holes and are also reported as an inquiline of social insects (S. latus by the ant Lasius brunneus). The genus Trochoideus is poorly studied ecologically. The most common and widespread species T. desjardinsi has been reported feeding on Auricularia sp. (Auriculaceaea), and could be myrmecophilous (associated with Plagiolepis longipes) or termitophilous (associated with Eutermes ceylonicus and Termes gilvus).
For the fossil species described in the present paper, a similar biology (subcortical or inquiline in moist forest habitats with numerous over-matured trees, that are infested by fungi, and have sources of rotten wood with an abundance of different termite and ant species) is assumed. The termite genus Termopsis Heer, with two species described from Baltic amber (Engel et al., 2007), could be a potential host for the larvae of the newly described Trochoideus species. The larvae of the newly described Symbiotes could be associated with the numerous and different Lasius s. str. species known from the deposit, particularly with the most abundant species L. schiefferdeckeri Mayr (Dlussky, 2011).
The distribution of the handsome fungus beetles represented in Baltic amber is also interesting. A connection between the Eocene fauna of Europe and recent taxa in Asia is a common biogeographic pattern seen among Baltic amber fossils (Larsson, 1978; Engel, 2001). But such affinity is not unequivocally supported by the fossil endomychids. Some genera are wholly extinct, but have affinities to lineages today occurring in the Holarctic (e.g., Glesirhanis), or Oriental region (Laima gen. nov., and Palaecoryphus gen. nov.), are palaeoendemic and represent archaic lineages (Gramboale gen. nov.), or are taxonomically unclear (Phymaphoroides). Symbiotes has a Holarctic distribution. Trochoideus is widespread in the tropics around the world. Both of the extant genera found in Eocene Baltic amber co-occur today on the territory of the southern USA only. Other handsome fungus beetles reported from Baltic amber (Shockley and Alekseev, 2014) are Palaearctic (Hylaia Chevrolat, 1836, and Leiestes Chevrolat, 1836); distributed in the Holarctic, Ethiopian and Oriental regions (Mycetina Mulsant, 1846, and Lycoperdina Latreille, 1807); or are cosmopolitan (Mycetaea Stephens, 1829, and Holoparamecus Curtis, 1833). One pattern that may be worth exploring further is the concentration of relict taxa within peripheries of continental blocks: eastern and southern Asian coasts (from Japan to Vietnam); south-eastern and western coasts of North America; southern Mediterranean coasts of Europe. These areas with a non-arid subtropical climate may be the most likely places to search for surviving lineages from the Eocene beetles observed in European ambers.
ACKNOWLEDGMENTS
The authors are very thankful to C. and H.W. Hoffeins (Hamburg, Germany) for loan of the very interesting fossil material. We are also very grateful to P.I. Alekseev (St. Petersburg, Russia) for the excellent photographs of the specimens and to A. Ślipiński (Australian National Insect Collection, CSIRO, Canberra, Australia) for reading and correcting an early version of this manuscript. Three anonymous reviewers are greatly appreciated for their valuable comments and corrections.
REFERENCES
Arrow, G.J. 1922. Coleoptera, Erotylidae and Endomychidae, from the Seychelles, Chagos, and Amirantes Islands. Annals and Magazine of Natural History, 10:73-84. https://doi.org/10.1080/00222932208632743
Blackburn, T. 1895. Further notes on Australian Coleoptera, with descriptions of new genera and species. Transactions of the Royal Society of South Australia, 19:201-258.
Blatchley, W.S. 1910. Family XVII. Endomychidae. The handsome fungus beetles. p. 533-539. In: Blatchley, W.S. (ed.), An Illustrated Descriptive Catalogue of the Coleoptera or Beetles (exclusive of the Rhynchophora) Known to Occur in Indiana. The Nature Publishing Co., Indianapolis.
Buquet, J.B.L. 1840. Note sur le genre Trochoideus, et description d’une nouvelle espèce. Revue Zoologique, 1840:173-175.
Carpenter, F.M. 1992. Treatise on Invertebrate Paleontology. Part R, Arthropoda 4, Volume 4. Superclass Hexapoda. Geological Society of America, Boulder, Colorado, 1-655.
Casey, T.L. 1916. Some random studies among the Clavicornia. Endomychidae. p. 139-146. In: Casey, T.L. (ed.), Memoirs on the Coleoptera, PA, Lancaster.
Chevrolat, L.A.A. 1836. [new taxa]. p. 361-422. In Dejean, P.F.M.A. (ed.), Catalogue des Coléoptères de la Collection de M. le Comte Dejean. Meguignon-Marvis, Paris.
Csiki, E. 1902. Endomychidae novae. Termeszetrajzi Fuzetek, 25:352.
Csiki, E. 1909. Coleoptera nova in Museo Nationali Hungarico. Annales Historico-Naturales Musei Nationalis Hungarici, 7:340-343.
Curtis, J. 1833. Characters of some undescribed genera and species indicated in the ‘Guide to an Arrangement of British Insects. The Entomological Magazine, 1:186-187.
Dalman, J.W. 1825. Om insekter inneslutne i copal, jemte beskrifning på några deribland förekommande nya slägten och arter. Kongelige Vetenskaps-Academiens Handlingar, 46:375-410.
Dlussky, G.M. 2011. The ants of the genus Lasius Fabricius (Hymenoptera, Formicidae) from the Late Eocene European ambers. Vestnik zoologii, 45(3):14-27. https://doi.org/10.2478/v10058-011-0014-5
Dury, C. 1912. New Coleoptera from Cincinnati, Ohio. Journal of the Cincinnati Society of Natural History, 21:102-103.
Engel, M.S. 2001. A monograph of the Baltic amber bees and evolution of the Apoidea (Hymenoptera). Bulletin of the American Museum of Natural History, 259:1-192.
Engel, M.S., Grimaldi, D.A., and Krishna, K. 2007. A synopsis of Baltic amber termites (Isoptera). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie), 372:1-20.
Gerstaecker, A. 1858. Monographie der Endomychidae, einer Familie der Coleopteren, p. 1-433. In: Gerstaecker, A. (ed.), Entomographien. Abhandlungen in Bereich der Gliederthiere, mit besonderer Benutzung der Koeniglichen Entomologischen Sammlung zu Berlin. W. Engelmann, Leipzig.
Gorham, H.S. 1887. Revision of the Japanese species of the coleopterous family Endomychidae. Proceedings of the General Meetings for Scientific Business of the Zoological Society of London, 1887:642-653.
Gorham, H.S. 1887-1899. Biologia Centrali-Americana. Insecta. Coleoptera. Vol. VII. Erotylidae, Endomychidae, and Coccinellidae. Taylor and Francis, London.
Gorham, H.S. 1896. Viaggio di Leonardo Fea in Birmania e regioni vicini. LXIX. Languriidae, Erotylidae and Endomychidae. Annali del Museo Civico di Storia Naturale di Genova, Ser. 2, 16:257-302.
Guérin-Méneville, F.E. 1838. Sur le genre Trochoideus, Westwood. Revue et Magasin de Zoologie Pure et Appliquée (2e Série), 1838:22-24.
Guérin-Méneville, F.E. 1857a. Trochoideus goudotii, Guer.-Men. nouvelle espèce (et peut-être nouveau genre) de Coléoptères de Colombie. Revue et Magasin de Zoologie Pure et Appliquée (2), 9:191-192.
Guérin-Méneville, F.E. 1857b. Matériaux pour une Monographie des Coléoptères du groupe des Eumorphides, et plus spécialement du genre Eumorphus. Archives Entomologiques, 1:237-280.
Hampe, C. 1850. Beschreibung einiger neuen Käferarten. Stettiner Entomologische Zeitung, 2:346-358.
Hoffeins, H.W. 2001. On the preparation and conservation of amber inclusions in artificial resin. Polish Journal of Entomology, 70:215-219.
Jacquelin du Val, P.N.C. 1858. [new taxa]. p. 129-232. In Jacquelin du Val, P.N.C. (ed,), Manuel Entomologique. Genera des Coléoptères d'Europe Comprenant leur Classification en Familles Naturelles, la Description de Tous les Genres, les Tableaux Synoptiques Destinés à Faciliter L'étude, le Catalogue de Toutes les Espèces, de Nombreux Dessins au Trait de Caractères. Tome deuxime. A. Deyrolle, Paris.
Joly, L.J. and Bordon, C. 1996. Two new species of Trochoideus Buquet from Venezuela (Coleoptera, Endomychidae), with comments on the Neotropical species groups. American Museum Novitates, 3169:1-10.
Kirejtshuk, A.G. 2000. On origin and early evolution of the superfamily Cucujoidea (Coleoptera, Polyphaga), comments on the family Helotidae. The Kharkov Entomological Society Gazette, 8:8-38.
Kirejtshuk, A.G. and Nel, A. 2009. New genera and species of Cucujiformia (Coleoptera, Polyphaga) from lowermost Eocene French amber. Denisia, 26:103-118.
Kirsch, T. 1876. Beiträge zur Kenntnis der Peruanischen Käferfauna auf Dr. Abendroth's Sammlungen basirt. Deutsche Entomologische Zeitschrift, 20:6-144.
Klebs, R. 1910. Über Bernsteineinschlüsse im allgemeinen und die Coleopteren meiner Bernsteinssammlung. Schriften der Physikalisch-ökonomischen Gesellschaft zu Königsberg, 51:217-242.
Kolbe, H. 1897. Coleopteren. Die Käfer Deutsch-Ost-Afrikas. Verlag von Dietrich Reimer, Berlin.
Kolbe, H. 1910. Die Coleopterenfauna der Seychellen. Mitteilungen aus dem Zoologischen Museum, Berlin, 5:1-49.
Larsson, S.G. 1978. Baltic Amber - a Palaeobiological Study. Volume 1. Scandinavian Science Press Ltd., Klampenborg, Denmark.
Latreille, P.A. 1807. Genera Crustaceorum et Insectorum Secundum Ordinem Naturalem in Familia Disposita, Iconobis Exemplisque Plurimis Explicta. Tomus tertius. Amand Koenig, Parisii et Argentorati.
Lea, A.M. 1922. On Australian Coleoptera. Part IV. Records of the South Australian Museum, 2:271-308.
Leach, W.E. 1815. Entomology. p. 57-172. In: Brewster, D. (ed.), Edinburgh Encyclopedia, volume 9. Balfour, Edinburgh.
LeConte, J.L. 1878. Additional descriptions of new species. The Coleoptera of Florida. Proceedings of the American Philosophical Society, 17:353-472.
Lucas, P.H. 1846. Exploration scientifique de l'Algérie pendant les années 1840, 1841, 1842 publiée par ordre du Gouvernement et avec le concours d'une commission académique Sciences Physiques Zoologie. Historie Naturelle des Animaux Articulés. Cinquieme classe. Insectes. Premier Ordre. Les Coléoptères, 2:1-360.
Motschulsky, V.I. 1856. Voyages. Lettres de M. de Motschulsky a M. Menetries. No. 3. New York le 15 Juillet 1654. Études Entomologiques, 5:21-38.
Mulsant, E. 1846. Histoire Naturelle des Coléoptères de France. Sulcicolles-Securipalpes. Maison, Paris.
Panzer, G.W.F. 1795. Entomologia Germanica Exhibens Insecta per Germaniam indigena, Secundum Classes, Ordines, Genera, Species, Adiectis, Synonymis, Locis, Observationibus. I. Eleuterata. Cum Tabulis Aeneis. Deutschlands Insectenfaune oder Entomologisches Taschenbuch für das Jahr 1795. Apud Fulseckeri Haeredes, Norimbergae.
Perty, J.A.M. 1832. Fascicule 2. p. 61-124. In: de Spix, J.B. and Martius, C.F. Ph. (eds.), Delectus Animalium Articulatorum, Quae in Itinere per Brasiliam Annis MDCCCVII-MDCCCXX Jussu et Auspiciis Maximiliani Josephi I. Bavariae Regis Augustissimi Peracto Collegerunt. Accedit Dissertatio de Insectorum in America Meridionali Habitantium Vitae, Genere, Moribus et Distributione Geographica. J.A.M. Perty, Monachii.
Poinar, Jr. G.O. and Poinar, R. 2008. What Bugged the Dinosaurs? Insects, Disease, and Death in the Cretaceous. Princeton University Press, Princeton.
Ponomarenko, A.G. 2002. Superorder Scarabaeoidea Laicharting, 1781. Order Coleoptera Linne, 1758. The beetles, p. 164-176. In: Rasnitsyn, A.P. and Quicke, D.L.J. (eds.), History of Insects. Kluwer, Dordrecht.
Redtenbacher, L. 1849. Fauna Austriaca. Die Käfer. Nach der analytischen Methode bearbeitet. Carl Gerold, Wien.
Reiche, L.J. 1847. [new taxa]. In Ferret, P.V.A. and Galinier, J.G. (eds.), Voyage en Abyssinie, dans les provinces du Tigre, du Samen et de l’Ahmara. Entomologie. Volume 3. Paulin, Paris.
Reitter, E. 1881. Coleopterologische Ergebnisse einer Reise nach Croatien, Dalmatien und der Herzegowina im Jahre 1879. Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien, 30:201-228.
Robertson, J.A., Ślipiński, S.A., Moulton, M., Shockley, F.W., Giorgi, A., Lord, N.P., McKenna, D.D., Tomaszewska, W., Forrester, J., Miller, K. B., Whiting, M.F., and McHugh, J.V. 2015. Phylogeny and classification of Cucujoidea and the recognition of a new superfamily Coccinelloidea (Coleoptera: Cucujiformia). Systematic Entomology, 40:745-778. https://doi.org/10.1111/syen.12138
Sasaji, H. 1970. Notes on the Formosan Endomychidae, with descriptions of a new genus and several new species (Coleoptera). Etizenia, 43:1-25.
Sasaji, H. 1990. The family Mychothenidae of Japan (Coleoptera). Esakia, Special Issue, 1:65-75.
Shockley, F.W. 2006. Discolomopsis dominicana, a new genus and species of handsome fungus beetle (Coleoptera: Endomychidae) from Dominican Amber. Insecta Mundi, 20:1-4.
Shockley, F.W. 2010. Micropsephodes bahamaensis, a new species of Anamorphinae (Coleoptera: Cucujoidea: Endomychidae) from the Bahamas, with a key to the New World genera of Anamorphinae. Insecta Mundi, 112:1-11.
Shockley, F.W. and Alekseev, V.I. 2014. Glesirhanis bercioi, a new genus and species from Baltic amber (Coleoptera: Endomychidae: Leiestinae) with a checklist and nomenclatural notes regarding fossil Endomychidae. Zootaxa, 3755(4):391-400. https://doi.org/10.11646/zootaxa.3755.4.5
Shoсkley, F.W. and Tomaszewska, K.W. 2007. First larval description for Symbiotes gibberosus (Lucas) (Coleoptera: Endomychidae). Annales Zoologici, 57(4):751-755.
Shockley, F.W., Tomaszewska, K.W., and McHugh, J.V. 2009a. An annotated checklist of the handsome fungus beetles of the world (Coleoptera: Cucujoidea: Endomychidae). Zootaxa, 1999:1-113.
Shockley, F.W., Tomaszewska, K.W., and McHugh, J.V. 2009b. Review of the natural history of the handsome fungus beetles (Coleoptera: Cucujoidea: Endomychidae). Insecta Mundi, 72:1-24.
Spahr, U. 1981. Systematischer Katalog der Bernstein- und Kopal-Käfer (Coleoptera). Stuttgarter Beiträge zur Naturkunde. Serie B (Geologie und Paläontologie), 80:1-107.
Stephens, J.F. 1829. A Systematic Catalogue of British Insects: being an Attempt to arrange all the hitherto discovered Indigenous Insects in Accordance with their Natural Affinities. Baldwin and Cradock, London.
Strohecker, H.F. 1943. Some fungus beetles of the family Endomychidae in the United States National Museum, mostly from Latin America and the Philippine Islands. Proceedings of the U.S. National Museum, 93:381-392.
Strohecker, H.F. 1953. Coleoptera Fam. Endomychidae, p. 1-140. In Wytsman P. (ed.), Genera Insectorum. Desmet-Verneuil, Bruxelles.
Strohecker, H.F. 1958. A synopsis of Philippine Endomychidae (Coleoptera). Fieldiana, Zoology, 42:19-48. https://doi.org/10.5962/bhl.title.3236
Strohecker, H.F. 1959. Les Endomychides Malgaches et les Trochoideites [Notes Synonymiques, Nouveautés]. Le Naturaliste Malgache, 11:77-85.
Strohecker, H.F. 1978. The American species of Trochoideus (Coleoptera: Endomychidae). The Coleopterists Bulletin, 32:349-354.
Strohecker, H.F. 1980. Eine neue Trochoideus -Art mit überblick über die asiatischen Formen (Col. Endomychidae). Deutsche Entomologische Zeitschrift, 27:89-92.
Thomson, C.G. 1863. Skandinaviens Coleoptera, synoptiskt bearbetade. Vol. 5. Tryckt uti Berlingska Boktryckeriet, Lund.
Thomson, J. 1860. Musée Scientifique ou recueill d’histoire naturelle, Heft 1. Chez l'auteur, Paris.
Tomaszewska, W. 2000. Morphology, phylogeny and classification of adult Endomychidae (Coleoptera: Cucujoidea). Annales Zoologici, 50:449-558.
Tomaszewska, W. 2004. A review of the genus Erotendomychus Lea (Coleoptera: Endomychidae). Annales de la Société Entomologique de France, 40:395-419. https://doi.org/10.1080/00379271.2004.10697430
Tomaszewska, W. 2005. Phylogeny and generic classification of the subfamily Lycoperdininae with re-analysis of the family Endomychidae (Coleoptera: Cucujoidea). Annales Zoologici, 55(supplement 1):1-172.
Tomaszewska, W. 2010. Endomychidae Leach, 1815, p. 442-454. In Leschen, R.A.B., Beutel, R.G., and Lawrence, J.F. (eds.), Handbook of Zoology, Vol. 2, Coleoptera. Walter de Gruyter GmbH and Co. KG, Berlin and New York.
Walton, L.B. 1912. Symbiotes duryi, a new species of Endomychidae. The Ohio Naturalist, 12:461-464.
Wasmann, E. 1922. Die Paussiden des zoologischen Staatsinstituts und zoologischen Museums zu Hamburg (ausser der Gattung Arthropterus) mit einem Anhang über die Endomychidengattung Trochoideus. Entomologische Mitteilungen aus dem Zoologisches Staatsinstitut und Zoologischen Museum in Hamburg, 39:12-38.
Westwood, J.O. 1833. On the Paussidae, a family of coleopterous insects. Transactions of the Linnean Society of London, 16:607-684.
Westwood, J.O. 1838. Descriptions of some new or but imperfectly known species belonging to the coleopterous family Paussidae. Transactions of the Entomological Society of London, 2:84-98.
