|
||||
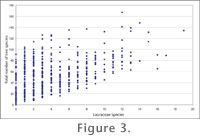
DISCUSSIONThe fossil record clearly includes additional genera, to the two which exist in New Zealand today, Beilschmiedia and Litsea. The identification of Endiandra is the first record in New Zealand. The case for Cryptocarya is confirmed, though this was fairly clear from the impressions of Holden (1982). The great diversity of the remaining cuticle suggests other genera were almost certainly present, perhaps ones now globally extinct. Conversely, neither of the two extant New Zealand genera is unequivocally present as fossils. Notable too, is that one extant genus in Australia with distinctive cuticle, Cinnamomum, has not been found as fossils in these deposits, and also some of the more distinctive species groups found today in Australia (for example the Endiandra jonesii group, Christophel and Rowett 1996). The current mean annual temperature for St Bathans (about 550 m above sea level) is in the order of 8°C (recording station at Ranfurly, 40 km to the east; this and following temperatures courtesy of Department of Geography, University of Otago, and NIWA, online). However, this depressed temperature reflects the mountainous and continental situation and a more realistic comparison with the low-lying topography of the Early Miocene would be coastal stations at equivalent latitudes. For instance, Oamaru Airport has a mean annual temperature of 10.6°C. In Southland, Invercargill Airport has a mean annual temperature of 9.7°C. Given the similar temperatures that Lauraceae extend to on tropical mountains, it is likely to be winter minima which are limiting Lauraceae in southern New Zealand today. Given the high diversity of the fossils, it seems clear that mean annual temperatures at the fossil localities were warmer than what they are now (southern New Zealand was at higher latitudes in the early Miocene, around 50-51°S; Lawver and Gahagan 2003). In broad terms, annual temperatures were probably similar to the rainforests of southeastern Queensland today, around 12-18°C (Hijmans et. al 2005). The presence of Lauraceae places a lower limit on warmth and usually is an indication of moist conditions. For instance the southern limit in New Zealand today is just south of Cook Straight, at about 42°S (Wardle 1991), where mean annual temperatures are about 12°C (Department of Geography, University of Otago, and NIWA, online). The altitudinal limit of Beilschmiedia tawa at 41S is about 1500 m (Wardle 1964). In Australia the southern limit of broad-leaved Lauraceae is the northern edge of Bass Straight at around 39°S (Cameron 1987; Helman 1987; Mills 1987; the leafless Cassytha is present in Tasmania) at about 14.5°C (Hijmans et. al 2005). In South Africa Lauraceae extend along the wettest fringe to the southernmost part of the country (Palgrave 1995) and in South America to about 40-41S and about 11C MAT (for example, species lists in Fraver et al. 1999; Veblen et al. 1979). In more tropical latitudes the altitudinal limits may reach cooler mean annual temperatures. For instance, the upper limit of Lauraceae on Mt Emei, China, is at about 8°C (Tang and Ohsawa 1997, 1999) and on Mt Kerinci, Sumatra (Ohsawa et al. 1985). Species diversity drops towards these southern boundaries and although only one or two species exist in these regions (for instance only three species occur in all of New Zealand today), they may dominate the biomass. In tropical rainforests Lauraceae tend to be much less prominent as biomass is shared amongst many more families. Lauraceae are typically components of a rainforest, and in Australia, with the exception of Cassytha, they are absent from vegetation with a fire-based ecology. In warmer areas as rainfall decreases, but fire remains absent, dry rainforests persist, but beyond some moisture limit, broad-leaved Lauraceae disappear. For instance, they are absent from the dry rainforest at Forty Mile Scrub, north Queensland, where mean annual temperature is about 23°C and mean annual rainfall is about 800 mm, but about 80% of this falls within a four-month period (Fensham 1996). More uncommonly, Lauraceae may be prominent in drier regions, for example in the sclerophyllous vegetation of Chile (Ramirez et al. 2004). The presence of a high diversity of Lauraceae most likely indicates wet, fire-free conditions. All the St Bathans samples considered here come from a stratigraphic level below indications of significant dry periods and below the evidence of widespread fire in the Manuherikia Group (Pole and Douglas 1998; Pole 2003).
|
||||
|
|