

 Ademosynidae (Insecta: Coleoptera): A new concept for a coleopteran key taxon and its phylogenetic affinities to the extant suborders
Ademosynidae (Insecta: Coleoptera): A new concept for a coleopteran key taxon and its phylogenetic affinities to the extant suborders
Article number: 20.2.31A
https://doi.org/10.26879/739
Copyright Palaeontological Association, June 2017
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 28 October 2016. Acceptance: 1 June 2017
{flike id=1896}
ABSTRACT
†Ademosynidae is a controversial extinct family of Coleoptera, first discovered in Australian Upper-Triassic deposits. The last known representative is known from the Early Jurassic of Kirghizstan. Type material from the collection of Moscow Paleontological Institute is re-described here and interpreted phylogenetically. The results show that the family as presently defined is not monophyletic, but possibly divided among two of four extant coleopteran suborders. We establish a new narrower concept of the family including the genera Ademosyne Handlirsch, 1906, Dolichosyne Ponomarenko, 1969, Gnathosyne Ponomarenko, 1969, Cephalosyne Ponomarenko, 1969, Petrosyne Ponomarenko, 1969, and Sphaerosyne Ponomarenko, 1969. This monophyletic unit is characterized by two synapomorphies of the prothorax, the absence of anterolateral pronotal angles and a rounded protruding anterior pronotal margin covering the posterior part of the head. Additional characters are large mandibles distinctly protruding anteriorly, striated elytra, an indistinct pronoto-elytral angle resulting in a stream-lined body, and a cuticular surface with coarse punctures. Based on the lack of the two prothoracic apomorphies, the Permian Archosyne Ponomarenko et al., 2014 is excluded from the family. As its placement remains very uncertain, it should be treated as Coleoptera incertae sedis. Ranis Ponomarenko, 1968 also lacks the pronotal features defining †Ademosynidae. Based on the internalized propleuron it is transferred to the megadiverse suborder Polyphaga. In its evolutionary history in the Mesozoic, †Ademosynidae reflects a general trend in Coleoptera, an increasing reinforcement of the configuration of the thoracic sclerites.
Evgeny V. Yan. Institut für Spezielle Zoologie und Evolutionsbiologie, Friedrich Schiller University Jena, 07737, Germany and Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya str. 123, Moscow, 117997 Russia. yan-e@mail.ru
Rolf G. Beutel. Institut für Spezielle Zoologie und Evolutionsbiologie, Friedrich Schiller University Jena, 07737, Germany. rolf.beutel@uni-jena.de
Alexander G. Ponomarenko. Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya str. 123, Moscow, 117997 Russia. aponom@paleo.ru
Keywords: Coleoptera; †Ademosynidae; paleontological reconstruction; early evolution
Final citation: Yan, Evgeny V., Beutel, Rolf G., and Ponomarenko, Alexander G. 2017. Ademosynidae (Insecta: Coleoptera): A new concept for a coleopteran key taxon and its phylogenetic affinities to the extant suborders. Palaeontologia Electronica 20.2.31A: 1-22. https://doi.org/10.26879/739
palaeo-electronica.org/content/2017/1896-redescription-of-ademosynidae
INTRODUCTION
†Ademosynidae is an extinct coleopteran family of small oval beetles, rarely longer than 1 cm. Six genera and 13 species were in the classical treatment of extinct and extant Archostemata of Ponomarenko (1969). All described ademosynids were found in Mesozoic localities of Early Triassic - Early Cretaceous age in Kirghizia, Kazakhstan, Russia, Argentina, and Australia. The family described more than 100 years ago by A. Handlirsch (1906) is still the subject of intensive phylogenetic dispute (e.g., Ponomarenko, 1969; Beutel, 1997; Beutel et al., 2008): they could either be considered as early representatives of the most advanced and diverse beetle suborder Polyphaga, or unusual members of the relict taxon Archostemata. The latter group is closest to the most ancestral extinct beetles in terms of morphological features and probably also life habits (Ponomarenko, 1969; Crowson, 1975, 1981; Lawrence and Newton, 1982; Lawrence, 1999; Beutel et al., 2008; Hörnschemeyer, 2016; Kirejtshuk et al., 2016). Archostemata was placed as the sistergroup of the remaining extant coleopteran lineages (Beutel and Haas, 2000), but the systematic position is still controversial (e.g., Kukalová-Peck and Lawrence, 2004; Misof et al., 2014; McKenna et al., 2015).
Seemingly mixed characteristics from both suborders impede the phylogenetic interpretation of the accessible morphological features of †Ademosynidae. A conspicuous feature well visible in fossils are the generally striated elytra, similar to those of some groups of Polyphaga (e.g., Hydrophilidae part., Elateridae part.) and Adephaga (e.g., Gyrinidae part., Carabidae part.): elytral cells of †Ademosynidae were transformed into internal supporting structures, the columelli. Such elytral organization clearly differs from clathrate or lattice-like elytra of the archostematan Cupedidae and Ommatidae, also referred to as window-puncture pattern. However, lattice elytral organization is not characteristic for the entire Archostemata, since species of Crowsoniellidae, Micromalthidae, and Jurodidae have glabrous or striated elytra. Other morphological features found in most ademosynids are reduced paranotalia and highly movable, unfused abdominal ventrites, similar to those of Scirtoidea. In dorsal view, they superficially resemble beetles of the families Hydrophilidae, Derodontidae, Eucnemidae, Throscidae, or Scirtidae. However, the presence of external propleurites and notopleural sutures, mesocoxal cavities partially closed by metanepisterna, and external metatrochantins are features typical of Archostemata (Friedrich et al., 2008).
In the present study we re-describe ademosynid fossils and evaluate the accessible morphological features phylogenetically, based on older and more recent studies on the morphology of extant and extinct beetles (e.g., Ponomarenko, 1969, 2004; Beutel et al., 2008; Friedrich et al., 2008; Yan et al., 2013). The main aim is to establish a well-defined monophyletic ademosynid unit, but the phylogenetic affinities are also discussed including a numerical phylogenetic evaluation of a modified character set from a previous study on a newly erected family †Peltosynidae (Yan et al., 2017). This may contribute to a better understanding of the basal splitting events in the Permian-Triassic boundary zone and the earliest evolution of the forming suborders of crown-group Coleoptera.
†Ademosynidae Research History. The genus Ademosyne as described by Handlirsch (1906) comprised only the two species, A. major Handlirsch, 1906 and A. minor Handlirsch, 1906, both from the Australian Upper Triassic locality Denmark-Hill. They were placed in the polyphagan family Hydrophilidae (Staphyliniformia) with mostly aquatic extant representatives. An almost completely preserved specimen of A. major is 4.5 mm long and shows mainly details of the dorsal aspect: pronotum, scutellar shield, and a pair of striate elytra. Among characters of the ventral side, the mesocoxal cavities and four abdominal sternites could be observed. A. minor was only represented by elytra of about 3 mm length. In subsequent years many isolated elytra from the type locality were described as new Ademosyne species: eight by Tillyard and Dunstan (1916), nine by Tillyard and Dunstan (1923), and a single species by Rohdendorf (1961) from the Upper Permian of the Kuznetsk Basin. Rohdendorf (1961) was the first who transferred Ademosyne from Polyphaga into Archostemata incertae sedis. Ponomarenko (1968) extended Rohdendorf’s concept by creating the separate archostematan family †Ademosynidae with two genera: Ademosyne (only the type species A. major) and a newly described monotypic genus Ranis Ponomarenko, 1968 from the Upper Jurassic of Kazakhstan. The rest of Ademosyne, known only as separate elytra, was transferred to another family by Ponomarenko (1968), †Permosynidae (Archostemata), which was erected by Tillyard in 1924. In the following year, the taxonomic concept of †Ademosynidae was greatly expanded by adding five genera and 11 species (Ponomarenko, 1969): three species of Dolichosyne Ponomarenko, 1969, three species of Ademosyne, and the monotypic genera Cephalosyne Ponomarenko, 1969 and Petrosyne Ponomarenko, 1969 reported from the Lower-Middle Triassic of Kirghizia. Additionally, the monotypic genus Gnathosyne Ponomarenko, 1969 was discovered in Lower Jurassic deposits of Eastern Kazakhstan. The youngest genus Sphaerosyne Ponomarenko, 1969 was reported from the Lower Cretaceous Transbaikalian locality Baissa. Much later three additional new species of Ademosyne were found in the early Late Triassic of Argentina (Martins-Neto, Gallego and Mancuso, 2006), however all of them only as separated elytra and following Ponomarenko’s concept (1968) likely belonging to †Permosynidae. In 2009 abundant material was obtained from the Middle-Late Jurassic locality Daohugou in North-East China. Three newly described genera Lasiosyne Tan and Ren, 2009, Pappisyne Tan and Ren, 2009, and Brachysyne Tan and Ren, 2009 showed a hitherto unmatched level of structural details, and were considered as crucial for understanding the morphology, subordinal placement and phylogenetic relationships of †Ademosynidae according to the authors (Tan and Ren, 2009). However, these genera were soon re-identified as elateriform Polyphaga, excluded from †Ademosynidae (Kirejtshuk et al., 2010), and placed within the superfamily Byrrhoidea (Yan et al., 2013). In 2014 Archosyne Ponomarenko et al., 2014 was described from Middle-Late Permian deposits of Eastern China (Ponomarenko et al., 2014), the most ancient ademosynid beetle. However its placement in to Ademosynidae was not confirmed in the current study.
Despite a long research history of †Ademosynidae, the obviously unstable taxonomic placement, and the candidacy of belonging to the stem group of the megadiverse Polyphaga, no reviews or phylogenetic analyses were focused on the family. The main reasons impeding studies of †Ademosynidae are limited fossil material and the poor preservation quality. For a reliable subordinal assignment, a complete body impression is necessary, with at least the ventral side preserved. Only this type of preservation traces crucial features such as the prothoracic organization, the type of mesocoxal closure, the configuration of the ventral meso- and metathoracic sclerites, and details of the head and its appendages. This is now made possible with type material deposited in the Moscow Paleontological Institute, including species of the genera Ademosyne, Dolichosyne, Ranis, Cephalosyne, and Petrosyne.
GEOLOGICAL BACKGROUND AND MATERIAL
The material studied was collected in Kirghizia, Osh province, Batken region, Dzhaylyaucho, Middle-Upper Triassic, Madygen Formation and Eastern Kazakhstan, Saur Range, southern slope of Saikan mountain, right bank of Akkolka river, 300-400 m northwest from the ravine Toktyrbai-sai, Kenderlyk, Upper Triassic, Tologoy Formation.
Madygen Formation, Ladinian and/or Carnian according to Dobryskina (1980, 1982), is either of later Middle or earlier Late Triassic age. The deposit of southern Fergana Valley (an area of junction of Uzbekistan, Kirghizia, and Tadzhikistan) includes clay layers very rich in fossil insects. Kenderlyk deposits are insectiferous mudstones of the Tologoy Formation, associated with the Norian and Rhaetian of the Late Triassic (Dobruskina, 1980, 1982).
Petrosyne liassica Ponomarenko, 1969 was found in Early Jurassic mudstones of coal-bearing formation near Kyzyl-Kiya railway station, South Fergana, Kirghizstan (Martynov, 1937).
The holotype of Sphaerosyne Ponomarenko, 1969 was not found in PIN RAS collection. It was probably lost and could not be included in our study. The accessible specimens of other genera were examined dry using Leica M165C microscopes. The photographs were prepared using attached digital cameras Leica DFC. Line drawings were readjusted on photographs using image-editing software CorelDRAW and X4. Photographs were processed with Adobe Photoshop CS and CorelDRAW X4.
Drawing conventions are the following: solid line, distinct margin; dashed, indistinct, and structures overlapping each other; dashed and dotted, fold; light grey, cavities; dotted area, membranous connections. On the reconstruction figures: red lines are reconstructed parts, solid black lines – characters restored after type material. The following measurements were recorded (depending on the state of preservation): total body length (including length of everted abdominal sternites and genitalia), body width; length and width of elytra, head, pronotum, and abdomen; for legs only the length was specified; tarsal length excludes length of claws (see Table 1). All fossils except Archosyne permiana Ponomarenko et al., 2014 and type species of Ademosyne major Handlirsch, 1906 are deposited in the Paleontological Institute, Russian Academy of Sciences (PIN RAS).
In the present study we use the concept of Archostemata of Beutel et al. (2008), with families †Tshecardocoleidae, †Permocupedidae, †Rhombocoleidae, and †Triadocupedidae regarded as stem group coleopterans.
References for the comparative morphology reconstruction are: Stenocyphon sasaji Lawrence, 2001 after Lawrence (2001), figures 1-4, pp. 353-354, with modifications; Epiphanis cornutus (Eschscholtz, 1829) redrawn using reference photos from www.biodiversity.ubk.ca/entomology/main/coleoptera/Eucnemidae; Dolichosyne sulcata Ponomarenko, 1969 and Gnathosyne akkolkensis Ponomarenko, 1969 reconstructed specimens available for present study; Archosyne permiana Ponomarenko et al, 2014 restored from the original publication, phototable 13.5a-d and figure 3, p. 195; †Taldycupedidae representative is reconstructed using Tecticupes heckeri, Rohdendorf, 1961 (Ponomarenko,1969), figure 68a, b, p. 122, with modifications; in †Permocupedidae reconstruction of head, pronotum, elytra, and ventral aspect based on Archicupes reichardii, Rohdendorf, 1961 (Ponomarenko,1969), figure 41, p. 69, with modifications, antennae drawn with reference to Permocupes sojanensis Ponomarenko, 1963 (Ponomarenko, 1969), figure 37, p. 65
Archostematan (Prolixocupes latreillei [Solier, 1849]) for strict consensus tree is redrawn from Hörnschemeyer (2016), figure 5.3 A, B, p. 45 with minor changes; †Peltosyne is copied from Yan et al. (in press); polyphagan beetle (Bolboceras floridensis [Wallis, 1928]) redrawn from Scholtz and Grebennikov (2016).
Phylogenetic analyses were based on 36 coded characters (Appendix 1) mostly from Yan et al. (in press), with a main focus on features that can be observed in well-preserved fossils (e.g., shape of pronotum, configuration of ventral thoracic sclerites). The taxon sampling included two megalopteran outgroups (Sialis, Chauliodes) and 28 coleopteran ingroup taxa representing all four extant suborders and the coleopteran stemgroup (14 extant, 14 extinct). Parsimony analyses were carried out with NONA (Goloboff, 1995) (ratchet, 1000 replicates). All characters were equally weighed and unordered.
SYSTEMATIC PALEONTOLOGY
Order COLEOPTERA Linnaeus, 1758
Suborder ARCHOSTEMATA Kolbe, 1908
Family †ADEMOSYNIDAE Ponomarenko, 1968
Ademosynidae: Ponomarenko, 1968, p. 128
Type genus. Ademosyne Handlirsch, 1906.
Type species. Ademosyne major Handlirsch, 1906.
Genera included .Dolichosyne Ponomarenko, 1969; Gnathosyne Ponomarenko, 1969; Ademosyne Handlirsch, 1906; Cephalosyne Ponomarenko, 1969, Sphaerosyne Ponomarenko, 1969, Petrosyne Ponomarenko, 1969.
Differential diagnosis. Differs from almost all extant and extinct groups of beetles by the pronotum without anterior angles and with a rounded anterior margin. Differs from Archostemata (except for Jurodidae) and stemgroup Coleoptera (except for †Rhombocoleidae) in elytra with punctate stria.
Revised description. Small to medium sized, oval beetles. Body surface usually covered with very small dense punctures, without large rounded or pentagonal tubercles or scale-like setae.
Head prognathous, large, transverse, almost quadrangular; half as wide as pronotum, more than 0.3 of its length concealed by anterior pronotal projection; dorsal protuberances absent, and also ventral antennal grooves and constricted neck region. Mandibles long and massive, distinctly protruding forward, at least half as long as head capsule, weakly curved apically; mandibular denticles missing. Compound eyes medium sized to large, not protruding laterally; ocelli absent.
Pronotum more or less distinctly narrowing anteriad; pronotal disk strongly convex and rounded anteriorly; anterolateral angles absent; propleura usually exposed, narrow, strip-like, tapering anteriorly. Well-developed prosternal process separating procoxae usually reaches posterior procoxal margin or slightly beyond it, rarely only half as long as procoxae, often widening apically; procoxal cavities open posteriorly.
Elytra with 9-12 punctate striae, not or only slightly protruding beyond abdominal apex posteriorly.
Mesoventrite very short, with distinct longitudinal suture; mesoventral pit almost always well developed; transverse suture absent. Mesocoxae distinctly separated. Meso- and metaventrite closely adjacent; externally visible membranous articulatory area absent.
Metaventrite trapeziform, with longitudinal and transverse sutures, the latter in some cases partially obliterated. All trochantins externally visible but usually with a narrow exposed portion. Metacoxae laterally reaching beyond lateral margin of metaventrite; coxal plates absent, posterior coxal margin only slightly excavated. Legs cursorial, with five unmodified tarsomeres.
Remarks. The re-examination of ademosynid type material revealed that the propleura is quite narrow and tapers anteriorly (internalized in Petrosyne). The presence of a mesoventral transverse suture was not confirmed for the family. These two characters differ from the original descriptions (Ponomarenko, 1969).
For key to genera of †Ademosynidae see Table 2.
Genus DOLICHOSYNE Ponomarenko, 1969
Dolichosyne: Ponomarenko, 1969, p. 126
Type species.Dolichosyne confragosa Ponomarenko, 1969; by original designation.
Differential diagnosis. Protrochantins more or less rectangular, length and width equal; apex of prosternal intercoxal process with lateral protrusions (if not, then mandibles with distinctly curved apices and pronotum with median longitudinal depression); metacoxal width two times exceeding length, coxae with maximum length medially.
Revised description. Head prognathous, large, at least 0.3 shorter than pronotum, rectangular, with pronotum covering at least basal fourth; moderately retracted into prothorax; subocular sutures on ventral side present. Pronotum with lateral bead; pronotal disk strongly convex. Posterior pronotal margin strongly bisinuate, paranotalia triangular. Anterior prosternal margin located distinctly posterior to anterior margin of pronotum, distance about fourth of pronotal length. Prosternum as long as globular, widely separated procoxae. Prosternal process widened at apex, in some cases with pair of lateral protrusions. Scutellar shield small, ovoid.
Species included. Dolichosyne confragosa; D. rostrate; D. sulcata.
Dolichosyne confragosa Ponomarenko, 1969
D. confragosa: Ponomarenko, 1969, p. 127
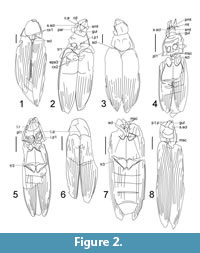
Figure 1.1-1.3, Figure 2.1-2.3
 Material. Holotype: PIN 2069/1336, strongly damaged. Middle Asia, Dzhailyaucho, Lower Triassic. Paratypes: PIN 1725/925, 2069/1340, 2070/1551 from the same locality.
Material. Holotype: PIN 2069/1336, strongly damaged. Middle Asia, Dzhailyaucho, Lower Triassic. Paratypes: PIN 1725/925, 2069/1340, 2070/1551 from the same locality.
Revised description. Mandibles 0.6 as long as head, distal third roundly curved and acute. Eyes large (Figure 2.2, c.e), as long as parietals (Figure 2.2, par), inner margins rounded, close to mandibular bases. Gular plate parallel-sided, narrow (Figure 2.2, gul), about 0.3 as wide as maximum width of submentum (Figure 2.2, smt). Submentum subtrapezoidal, as long as eye, with small lateral incisions on anterior angles for cardines (Figure 2.2, cd). Posterior angles of pronotum not protruding posterolaterally. Protrochanters 0.3 of profemoral length (Figure 2.2, tr1). Prosternal intercoxal process parallel-sided (Figure 2.2, i.p1), with apex abruptly widening apically and forming distinct lateral projections. Scutellar shield 0.6 as long as prosternal process (Figure 2.2, scl). Elytra with 11 striae, 2nd and 3rd striae (counting from sutural margin) shortened, half as long as elytra, 6th and 8th merging near elytral apex enclosing 7th, not reaching elytral apex. See measurements in Table 1.
In contrast to other species of Dolichosyne gular plate is only one third as wide as submentum; eyes large, longer than half of mandible.
Remarks. Due to strong damage the holotype it is not shown in Figure 1-Figure 2. The paratype 1725/925 is lost.
Dolichosyne rostrata Ponomarenko, 1969
D. rostrata: Ponomarenko, 1969, p. 129
Figure 1.4, Figure 2.4
 Material. Holotype: PIN 2555/1735, beetle without legs and antennae; Middle Asia, Dzhailyaucho, Lower Triassic.
Material. Holotype: PIN 2555/1735, beetle without legs and antennae; Middle Asia, Dzhailyaucho, Lower Triassic.
Revised description. Mandibles longer than half of head, only very slightly curved apically. Eyes relatively small, half as long as parietals, elongate-oval, not protruding laterally. Gular plate half as wide as head, submentum rectangular, same size as gular plate (Figure 2.4, gul, smt). Mentum small, transverse, half as long as submentum, anterior margin weakly rounded; prementum as long as mentum, half as wide, with pair of distinct cavities for labial palpi (Figure 2.4, mt, pmt). Pronotum with posterior angles distinctly protruding posterolaterally. Propleurites triangular (Figure 2.4, pl1). Prosternal intercoxal process distinctly widening posteriorly, with abruptly widened apex forming small lateral projections. Metaventrite with anterior angles strongly protruding anteriad; anterior intercoxal process with wide blunt apex. Elytra with at least eight striae. See measurements in Table 1.
Eyes smaller than in other species, less than half as long as mandibles, separated from mandibular bases at distance equal to their length; posterior pronotal angels distinct, projecting posterolaterally; anterior metaventral process with wide blunt apex.
Dolichosyne sulcata Ponomarenko, 1969
D. sulcata: Ponomarenko, 1969, p. 128
Figure 1.5-1.8, Figure 2.5-2.8
Material. Holotype: PIN 2240/274, beetle without anterior half of head and without legs; Middle Asia, Dzhailyaucho, Lower Triassic. Paratypes: PIN 2240/2250, 2240/191, 2240/234, 2240/305 from same locality.
PIN 2240/2250, 2240/191, 2240/234, 2240/305 from same locality.
Revised description. Distal third of mandible distinctly curved. Eyes half as long as mandibles. Head capsule with pair of large supraantennal tubercules located near anterior margin of frons and separated by longitudinal ridges (Figure 2.5, l.r). Gular plate long, about one third of pronotal length, parallel-sided, with distinct posterior tentorial pits. Pronotum with longitudinal median depression (Figure 2.5, l.d). Propleurites straight on outer sides; intercoxal process as long as procoxae, slightly widening apically (Figure 2.5, pl1, i.p1). See measurements in Table 1.
Body more strongly elongated than in other species of the genus, four times as long as wide; mandibular apices more strongly curved; prosternal process only slightly widening apically, lateral projections absent.
Remarks. Pronotum and prosternum of holotype shifted from each other, thus appearing asymmetrical.
Genus GNATHOSYNE Ponomarenko, 1969
Gnathosyne: Ponomarenko, 1969, p. 129
Type species. Gnathosyne akkolkensis Ponomarenko, 1969 by original designation.
Differential diagnosis. Anterior prosternal margin almost straight, very slightly concave; metacoxae three times as wide as long, not narrowing laterally.
Revised diagnosis. Head transverse, distinctly wider than long (not counting mandibular length). Mandibles closely spaced. Antennal insertions dorsal. Eyes large, ovoid, inserted laterally. Pronotum transverse, only slightly narrowing anteriorly; posterior angles very small and acute. Head strongly retracted below pronotum. Anterior prosternal margin distinctly posterior to anterior margin of pronotum, distance about one third of pronotal length. Prosternal intercoxal process not projecting beyond procoxae. Propleurites only slightly narrower anteriorly. Mesoventrite transverse, almost quadrangular, with very short anterior process and small rhomboid cavity. Metaventrite with very long anterior intercoxal process; longitudinal suture distinct only on basal half; paracoxal suture distinct.
Species included. Monotypic.
Gnathosyne akkolkensis Ponomarenko, 1969
G. akkolkensis: Ponomarenko, 1969, p. 129
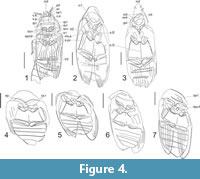
Figure 3.1, Figure 4.1
 Material. Holotype: PIN 2496/7, beetle lacking large parts of antennae, legs and abdominal apex; Eastern Kazakhstan, Saur mountain, Kenderlyk; Lower Jurassic, Tologoy Fm.
Material. Holotype: PIN 2496/7, beetle lacking large parts of antennae, legs and abdominal apex; Eastern Kazakhstan, Saur mountain, Kenderlyk; Lower Jurassic, Tologoy Fm.
Revised description. Head transverse, 1.5 times as wide as long. Mandibles very closely spaced (Figure 4, md). Eyes twice as long as parietals (Figure 4, c.e). Antennal insertions elevated, closer to inner ocular margins than to each other. Scape (Figure 4, sc) equal to lengths of quadrangular 2nd and 3rd antennomeres; 4th antennomer only slightly longer than scape; antennomeres 1-3 with fine longitudinal sulci.
Prosternal intercoxal process widens apically. Metaventrite as long as width at posterior margin, which is twice as wide as anterior margin. Mesoventral cavity small, rhomboid (Figure 4, ms.p). Pro- and mesofemora slightly thickened; tibia widened distally (Figure 4, fm1, tb1). Intercoxal process of first visible abdominal sternite weakly elevated. Ventral side of head and pronotum covered with small punctures merging into transverse wrinkles, rest of ventral body surface with more widely spaced small punctures. Most parts of dorsum with sparse punctures, almost appearing glabrous. See measurements in the Table 1.
Remarks. Here we interpret a pair of S-shaped lines close to the apex of the prosternal intercoxal process as fine sulci on the back side of the pronotum. An alternative interpretation as a propleural closure of the procoxal cavities appears less likely as the procoxal cavities are posteriorly more or less widely open in all other genera of †Ademosynidae.
Genus ADEMOSYNE Handlirsch, 1906
Ademosyne: Handlirsch, 1906, p. 402; Dunstan, 1924, p.11
 Type species. Ademosyne major Handlirsch, 1906
Type species. Ademosyne major Handlirsch, 1906
Differential diagnosis. Protrochantins transverse, more than 3 times as wide as long; metacoxae more than four times as wide as long, slightly narrowing laterally.
Revised diagnosis. Head strongly retracted below pronotum. Anterior prosternal margin distinctly posterior to anterior margin of pronotum, distance about one third of pronotal length. Pronotum with bead, lacking protruding posterior angles. Pro- and mesocoxae oblique, transverse, wider than long. Protrochantins elongated, more than three times as wide as long.
Anterior half of longitudinal suture of metaventrite often obliterated. Metacoxal width more than four times exceeding length; metacoxae weakly narrowing laterally.
Visible abdominal sternites telescopically retractable within each other; first and fifth ventrites longer than others.
Species included. Ademosyne major; A. kirgizica; A. bacca; A. elliptica.
Ademosyne kirgizica Ponomarenko, 1969
A. kirgizica: Ponomarenko, 1969, p. 132
Figure 3.2-3.4, Figure 4.2, 4.3
Material. Holotype: PIN 2096/1369, beetle without antennae and legs; Eastern Asia, Dzhailyaucho, Lower Triassic. Paratype: PIN 2555/1696 from the same locality.
Revised description. Gular plate as wide as short submentum; cardo rectangular (Figure 4.3, gul, smt, crd). Prosternum in front of procoxae slightly shorter than prosternal intercoxal process. Mesocoxae large, oblique, three times longer from anterolateral to posteromesal edge than space between them. Free mesotrochantin large. Metaventrite with bead at anterior edge; less than three times as long as its basal width; posteriorly 1.5 times wider than anteriorly. Prosternum and meso- and metaventrite covered with large punctures, rest of ventral side of body with small punctures. See measurements in Table 1.
Differs from other Ademosyne species in the strongly concave anterior prosternal margin, the elongate body (2.5 times as long as wide), the mesoventrite without an acute anterior process, a mesosternal longitudinal suture passing through the pit of the mesoventrite (Figure 4.2, sl.2), and a mesanepisternum with longitudinal ridges.
Ademosyne bacca Ponomarenko, 1969
A. bacca: Ponomarenko, 1969, p. 133
Figure 3.5, 3.6, Figure 4.4, 4.5
Material. Holotype: PIN 2240/310, beetle without antennae and legs; Eastern Asia, Dzhailyaucho, Lower Triassic. Paratype: PIN 2240/294 from the same locality.
Revised description. Pronotum very short, posterior margin weakly bisinuated. Mesocoxae twice as wide as long, two times broader than space between them. Metaventrite strongly transverse, half as wide as maximum length at posterior margin, which is twice as wide as anterior margin; paracoxal suture straight, mesal portions of metacoxae weakly shifted posteriorly. Fifth abdominal ventrite 1.5 times longer than penultimate. Ventral side of body covered with small punctures. See measurements in Table 1.
Differs from other species in the shape of the pronotum, which is twice as wide as long; presence of very deep transverse procoxal rests (Figure 4.4, cx.r) of mesoventrite; metaventral paracoxal suture strongly shifted towards posterior margin.
Ademosyne ellyptica Ponomarenko, 1969
A. ellyptica: Ponomarenko, 1969, p. 133
Figure 3.7, Figure 4.6
Material. Holotype: PIN 2240/224, beetle without antennae and legs; Eastern Asia, Dzhailyaucho, Lower Triassic.
Revised diagnosis. Prosternum longer than procoxae; prosternal process nearly rectangular, with wide blunt apex. Mesocoxae 0.6 as long as their maximum width, space between them equal to width. Metaventrite very broad, at posterior margin half as wide as maximum length; posterior margin 1.5 times wider than anterior margin; anterior process with straight apex. Last abdominal sternite only slightly longer than penultimate. Thoracic ventrites covered with large punctures and abdominal sternites with small tubercles. See measurements in Table 1.
Differs from other species in shorter and widely separated mesocoxae, and a broad anterior process of the first abdominal ventrite with a blunt apex.
Genus CEPHALOSYNE Ponomarenko, 1969
Cephalosyne: Ponomarenko, 1969, p. 134
Type species. Cephalosyne capitata Ponomarenko, 1969 by original designation.
Differential diagnosis. Prosternal process with acute apex, slightly longer than procoxae; anterior process of metaventrite more than two times shorter than mesocoxae.
Revised diagnosis. Small beetles, with elongate, ovoid body. Procoxae strongly transverse. Intercoxal process acute, narrowing apically, distinctly longer than prosternum in front it. Elytra with widely rounded apex, about three times as long as pronotum, with punctate striae. Mesocoxae widely separated, ovoid-transverse. Metaventrite rectangular, very weakly narrowing anteriorly; longitudinal suture distinct, paracoxal suture visible only near posterior angles. Metanepisterna with thick median longitudinal ridges. Visible abdominal sternites not overlapping; apical sternite twice as long as penultimate.
Species included. Monotypic.
Cephalosyne capitata Ponomarenko, 1969
C. capitata: Ponomarenko, 1969, p. 134
Figure 3.8, Figure 4.7
Material. Holotype: PIN 2240/302, beetle without legs; Eastern Asia, Dzhailyaucho, Lower Triassic. Paratypes: PIN 2069/1342, 2240/130, 2240/130, 2240/273, 2555/1732 from the same locality.
PIN 2069/1342, 2240/130, 2240/130, 2240/273, 2555/1732 from the same locality.
Revised description. At least anterior margin of pronotum with a fine bead. Prosternal intercoxal process with thickened outer margin, 1.5 times longer than prosternum in front of it. Space between mesocoxae slightly narrower than mesocoxal length. Metaventrite 1.5 times wider than long at posterior margin. Mesanepisterna with longitudinal ridges. Ventral side covered with rather large, dense tubercles, dorsal side with smaller sparse tubercles. See measurements in Table 1.
Differs from other species by narrow acute prosternal intercoxal process and short, broad intercoxal process of metaventrite.
Genus PETROSYNE Ponomarenko, 1969
Petrosyne: Ponomarenko, 1969, p. 135
Type species. Petrosyne liassica Ponomarenko, 1969 by original designation.
Differential diagnosis. Anterior margin of metanepisterna straight; anterior process of metaventrite elevated.
Revised diagnosis. Medium sized beetles with elongate body. Lateral margins of prosternum curved. Procoxal rests short and narrow. Mesoventral posterior process very narrow. Metaventrite parallel-sided, its posterior angles protruding posterolateraly. Anteromedian process of metaventrite elevated, incised apically. Metacoxae laterally extending to outer margins of metanepisterna. Anterior margin of metanepisterna straight.
Species included. Monotypic.
Petrosyne liassica Ponomarenko, 1969
P. liassica: Ponomarenko, 1969, p. 135
Figure 5.1, 5.2
 Material. Holotype: PIN 166/39, South Fergana, Kirghizstan, Kyzyl-Kiya, Early Jurassic.
Material. Holotype: PIN 166/39, South Fergana, Kirghizstan, Kyzyl-Kiya, Early Jurassic.
Revised description. Pronotum beaded; prosternal intercoxal process 1.5 times longer than procoxae. Mesosternal longitudinal suture reaches half-length of mesoventral pit length. Metaventrite and metanepisterna with thickened margins. Visible abdominal sternites 1-4 equal in length; apical sternite1.5 times longer than penultimate, incised at the apex. See measurements in the Table 1.
DISCUSSION
A primary aim of this study was a concept with monophyletic †Ademosynidae. The genera re-described here - Dolichosyne, Gnathosyne, Ademosyne, Cephalosyne, and Petrosyne - are characterized by small or relatively small size, an evenly oval, streamlined body without protruding eyes and pronoto-elytral angle (posterior pronotal angles of Dolichosyne rostrata and Gnathosyne akkolkensis weakly protruding posterolaterally), striated elytra, and a dorsal surface pattern with fine punctures. Characteristic and apomorphic features shared by these five taxa are the absence of anterolateral pronotal angles and a rounded anterior margin of the pronotum, covering the posterior part of the head. Such a condition is absent in potentially related extant and extinct beetles. Another potential synapomorphy is the large size of the mandibles, which are distinctly protruding anteriorly. These derived features, part of the newly defined diagnosis for †Ademosynidae, suggest that the genus †Ranis (Figure 5.4-5.6) should be excluded from the family. In contrast to the five genera treated here, they possess very distinct anterolateral pronotal angles and mandibles which are ventrally oriented. Their propleuron is internalized, which suggests that they should be transferred to Polyphaga incertae sedis. Affinities with Byrrhoidea are possible but not sufficiently supported presently. Other features distinguishing †Ranis from †Ademosynidae as defined here are the transverse head, the distinct pronoto-elytral angle, and the distally strongly widening protibiae.
The genus †Petrosyne displays all main characters of †Ademosynidae, especially the shape of the pronotum and also the general habitus. However, their propleuron are not visible externally, and the metanepisterna is shortened and not included in the mesocoxal closure. Both features suggest possible affinities with Polyphaga. However, mainly based on the very characteristic pronotal features we interpret †Petrosyne as an aberrant genus of †Ademosynidae, with an independently acquired internalized propleuron and a non-archostematan closure of the mesocoxal cavity.
The Permian Archosyne (Figure 6) is excluded from †Ademosynidae based on its distinctly protruding anterior pronotal angles, a straight anterior pronotal margin, a prosternal intercoxal process shorter than the procoxae themselves, contiguous mesocoxae, and flattened elytral bases. Paired median pronotal tubercles and supraantennal tubercles are structural affinities with stem group Coleoptera and Archostemata (e.g., Ponomarenko, 1969; Beutel et al., 2008). Striated elytra and a coarse punctuation are features shared with †Peltosynidae, †Ranis and †Ademosynidae (Yan et al., 2017). The cladistic analysis of 36 characters places Archosyne in an unresolved polytomy with terminals of crown group Coleoptera, without indicating phylogenetic affinities with †Ademosynidae or members of the extant suborders. The presently available characters are obviously not sufficient for a reliable placement of the genus.
The evolution of †Ademosynidae is apparently characterized by a tendency towards an advanced configuration of thoracic elements, possibly also linked with simplifications of the thoracic musculature (Beutel and Haas, 2000). Morphological transformations in the extinct family result in a number of features characterizing groups of the megadiverse Polyphaga or the entire suborder. This complex event of parallel evolution includes propleural internalization, development of a pro-mesosternal interlocking mechanism, prosternal shortening, metanepisterna not participating in the closure of the mesocoxal cavity, and a shift of the metaventral paracoxal suture towards the posterior margin of the sclerite. The most ancestral states are found in the Lower Triassic †Dolichosyne, probably representing the groundplan of the monophyletic family. The most advanced condition was observed in the Early Jurassic †Petrosyne, with a distinctly increased rigidity of the thorax.
The phylogenetic affinities of †Ademosynidae are apparently important in the context of the evolution of the major coleopteran lineages emerging in the Permian - Triassic transition zone. A serious problem in beetle phylogenetics are the unclarified relationships of the four extant suborders. Morphologists either suggested a pattern with Polyphaga basal followed by Archostemata as sistergroup of Adephaga + Myxophaga (Kukalová-Peck and Lawrence, 2004), based on wing characters, or alternatively based on a broad spectrum of characters of all body regions a basal position of Archostemata and a sistergroup relationship between Adephaga and Polyphaga + Myxophaga (Beutel and Haas, 2000; Beutel et al., 2008; Friedrich et al., 2008). The results of molecular studies ]vary greatly, with a pattern Archostemata + (Myxophaga + [Adephaga + Polyphaga]) in Caterino et al. (2002: 18S rRNA), (Archostemata + Myxophaga) + (Adephaga + Polyphaga) in Hunt et al. (2007: 16S, 18S rRNA, COI, 1900 spp.), Polyphaga + (Myxophaga + [Archostemata + Adephaga]) in Misof et al. (2014: transcriptomes) or Polyphaga + (Adephaga + [Archostemata + Myxophaga]) in McKenna et al. (2015: eight nuclear genes).
A second problem is the limited availability of morphological characters in the impression fossils compared to extant species. Structural details of the head (e.g., mouthparts) are often not visible or ambiguous, the wings are not preserved in most cases, and internal features (e.g., muscles) are generally not accessible. Due to these circumstances, a phylogenetic assignment of the taxa under consideration here is very difficult compared to extant coleopteran groups.
A major question associated with †Ademosynidae was already discussed by two eminent coleopterists, R.A. Crowson and J.F. Lawrence: are they the earliest and most ancestral lineage of the megadiverse Polyphaga, or are they rather a result of parallel evolution within the relict suborder Archostemata, with some characteristics resembling polyphagan features but without close phylogenetic relationship. Crowson (1981) interpreted †Ademosynidae as a polyphagan stem-group, as did Lawrence (1999), pointing out various similarities in the habitus between †Ademosynidae and the polyphagan family Scirtidae, which belongs to the most basal extant branch within the suborder (e.g., McKenna, 2015).
In contrast to this interpretation, Ponomarenko, (1983) considered †Ademosynidae as aberrant archostematan beetles (in the wider sense, i.e., including stem group Coleoptera; Ponomarenko, 1969), with a tendency towards a polyphagan level of organization. Similar processes are known in evolutionary history, when unrelated groups, for instance among angiosperm plants or “reptiles”, independently develop similar syndromes of characters (Ponomarenko, 1983, 2004, 2008, 2009). In this sense, †Ademosynidae were interpreted as a terminal stage of an “evolutionary sequence” †Permocupedidae - †Taldycupedidae - †Ademosynidae (Figure 6). One of the most notable results of this hypothesized evolutionary transition was the transformation of elytral cells into inner supporting structures - columelli.
The results presented here underline that †Ademosynidae as we define them differ in a considerable number of features from extinct or extant archostematan beetles (e.g., Ponomarenko, 1969; Beutel et al., 2008; Friedrich et al., 2008). This includes the evenly sclerotized and striated elytra, the lack of a tuberculate body sculpture and scales, the streamlined body outline without pronoto-elytral angle, and well-developed posterior pronotal angles. †Ademosynidae also distinctly differ from most recent and extinct archostematans in their more generalized head structure without constricted neck and postocular extensions. They also lack the typical labial configuration with an enlarged prementum and a reduced mentum (Tetraphalerus and Crowsoniella are also exceptions in Archostemata), and a fully developed gular plate is usually present. In contrast to most archostematans, the ventral aspect of the ademosynid body appears mechanically more rigid due to the strongly developed prosternal process (Figure 6.6, 6.8), often widening apically and associated with a large and deep anteromedian pit of the mesoventrite. However, in contrast to Cupedidae and Ommatidae, †Ademosynidae probably lacked a propleuro-mesepisternal locking mechanism (Friedrich et al., 2008).
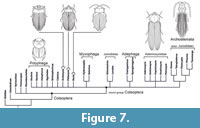 As pointed out by Lawrence (1999), some features of the ademosynid habitus (Figure 6.5-6.8) resemble conditions found in certain polyphagan beetles, especially of the series Scirtiformia (Figure 6.1, 6.2; Scirtidae: Stenocyphon Lawrence, 2001 and Nipponocyphon Lawrence and Yoshitomi, 2007), but also of Elateriformia (Figure 6.3, 6.4; Eucnemidae, Throscidae, and Artematopodidae) or Staphyliniformia (Hydrophilidae). However, closer scrutiny reveals that these characters are either unspecific, differ in structural details, or have apparently evolved several times independently (Figure 7). Independent evolution is very likely in the case of one of the most conspicuous ademosynid features, the striated elytra. A very similar condition is found in the extinct Peltosyne (Yan et al., 2017), in some groups of Adephaga (Gyrinidae part., Dytiscidae part., Carabidae part.), and in some isolated groups of Polyphaga (Hydrophilidea part., Elateridae part.). Similarly, a streamlined body form has doubtlessly evolved many times independently, often but not always correlated with aquatic habits.
As pointed out by Lawrence (1999), some features of the ademosynid habitus (Figure 6.5-6.8) resemble conditions found in certain polyphagan beetles, especially of the series Scirtiformia (Figure 6.1, 6.2; Scirtidae: Stenocyphon Lawrence, 2001 and Nipponocyphon Lawrence and Yoshitomi, 2007), but also of Elateriformia (Figure 6.3, 6.4; Eucnemidae, Throscidae, and Artematopodidae) or Staphyliniformia (Hydrophilidae). However, closer scrutiny reveals that these characters are either unspecific, differ in structural details, or have apparently evolved several times independently (Figure 7). Independent evolution is very likely in the case of one of the most conspicuous ademosynid features, the striated elytra. A very similar condition is found in the extinct Peltosyne (Yan et al., 2017), in some groups of Adephaga (Gyrinidae part., Dytiscidae part., Carabidae part.), and in some isolated groups of Polyphaga (Hydrophilidea part., Elateridae part.). Similarly, a streamlined body form has doubtlessly evolved many times independently, often but not always correlated with aquatic habits.
An ademosynid feature differing from almost all polyphagan groups is the relatively loose connection between the meso- and metaventrite. This also applies to Scirtoidea, in contrast to the rigid connection of these sclerites in almost all polyphagan lineages (and Myxophaga) (Beutel and Haas, 2000). However, in contrast to an exposed intersegmental membrane between the meso- and metaventrite in Scirtidae (Figure 6.2, mb.c), a specific connecting mechanism is present in †Ademosynidae: a distinctly developed acute anterior process of the metaventrite reaches the posterior process of the mesoventrite, which separates the mesocoxae.
Another character complex separating †Ademosynidae from Scirtiformia and many elateriform groups is the prognathous head, with massive, strongly protruding mandibles. A feature distinguishing †Ademosynidae from many basal elateriform taxa (Figure 5) (e.g., Dascillidae) is the shape of the procoxae. In †Ademosynidae they are usually more or less rounded (more elongated in Ademosyne and Cephalosyne), with almost equal length and width, in contrast to wider, transverse procoxae in different elateriform families (especially of the superfamily Byrrhoidea). This distinguishes them also from the recently established Late Triassic polyphagan family †Peltosynidae (Yan et al., 2017).
It is evident that a reliable placement of †Ademosynidae is not possible with the presently available characters (Figure 7; see also list of characters in Appendix 1). Features shared with other extinct or extant archostematan taxa are plesiomorphic, such as exposed propleurae (except for Petrosyne), at least partly external metatrochantins, and the type of mesocoxal closure including the metanepisternum. However, features linking †Ademosynidae with other coleopteran lineages including polyphagan subgroups are apparently based on convergency, and the cladistic analysis did not reveal phylogenetic affinities to any of the non-archostematan suborders. Apomorphies of Polyphaga are lacking in the groundplan of †Ademosynidae: the propleura is exposed in all genera except for Petrosyne and cervical sclerites (arguably plesiomorphic) are missing. Therefore, it appears appropriate to leave the family in Archostemata provisionally, in agreement with Ponomarenko (1969, 1983) and a cladistic study based on characters of extinct and extant members of this suborder (Beutel et al., 2008). A re-evaluation of newly discovered material with more visible details and possibly preserved hind wings may help to clarify the mosaic evolution of this extant group, with its intriguing mixture of features of different major lineages of Coleoptera.
ACKNOWLEDGMENTS
We thank anonymous reviewers for providing helpful suggestions. We are very grateful to R.G. Beattie for providing specimens of fossil beetles from the Australian Belmont locality and useful comments on this manuscript. This research was supported by an Alexander von Humboldt grant to EY and by Russian Foundation for Basic Research (16-04-01498).
REFERENCES
Baehr, M. 1975. Skelett und Muskulatur des Thorax von Priacma serrata Leconte (Coleoptera, Cupedidae). Zeitschrift für Morphologie der Morphologie der Tiere, 81:55-101. (In German)
Beutel, R.G. 1997. Über Phylogenese und Evolution der Coleoptera (Insecta), insbesondere der Adephaga. Verhandlungen des Naturwissenschaftlichen Vereins in Hamburg NF, 31:1-164. [in German]
Beutel, R.G. 1999. Phylogenetic analysis of Myxophaga (Coleoptera) with a redescription of Lepicerus horni (Lepiceridae). Zoologischer Anzeiger, 237:291-308.
Beutel, R.G., Ge, S.-Q., Hörnschemeyer, T. 2008. On the head morphology of Tetraphalerus, the phylogeny of Archostemata and the basal branching events in Coleoptera. Cladistics, 23:1-29.
Beutel, R.G., Haas, F. 2000. Phylogenetic relationships of the suborders of Coleoptera (Insecta). Cladistics, 16:103-141.
Caterino, M., Shull, V.L., Hammond, P.M., Vogler, A. 2002. Basal relationships of Coleoptera inferred from 18S rDNA sequences. Zoologica Scripta, 31:1-49.
Crowson, R.A. 1960. The phylogeny of Coleoptera. Annual Review of Ent omology, 5:111-134.
Crowson, R.A. 1975. The evolutionary history of Coleoptera, as documented by fossil and comparative evidence. Atti del X Congresso Nazionale Italiano di Entomologia, Sassari: 47-90.
Crowson, R.A. 1981. The Biology of Coleoptera. Academic Press, New York.
Dobruskina, I.A. 1980. Stratigraphic position of Triassic plant-bearing beds of Eurasia. Trudy Geologicheskogo Instituta Akademii Nauk SSSR, 3-161:346, Nauka, Moscow. (In Russian)
Dobruskina, I.A. 1982. Triassic Floras of Eurasia. Trudy Geologicheskogo Instituta Akademii Nauk SSSR, Nauka, Moscow. (In Russian)
Dunstan, B. 1924. Mesozoic insects of Queensland. Pt. 1. Introduction and Coleoptera. Queensland Geological Survey Publication, 273:1-74.
Eschscholtz, J.F. 1829. Elaterites, Eintheilung derselben in Gattungen. Entomologisches Archiv (Thon), 2(1):31-35. (In German)
Friedrich, F., Farrell, B.D., and Beutel, R.G. 2008. The thoracic morphology of Archostemata and the relationships of the extant suborders of Coleoptera (Hexapoda). Cladistics, 25:1-37.
Goloboff, P.A. 1995. NONA. Version 2.0. Available from: www.cladistics.com/.
Handlirsch, A. 1906-1908. Die fossilen Inscecten und die Phylogenie der rezenten Formen. Verlag von Wilhelm Engelmann, Leipzig. (In German)
Hlavac, T.F. 1972. The prothorax of Coleoptera: origin, major features of variation. Psyche, 79:123-149.
Hlavac, T.F. 1975. The prothorax of Coleoptera: (except Bostrichiformia - Cucujiformia). Bulletin of the Museum of Comparative Zoology, 147:137-183.
Hörnschemeyer, T. 2016. Archostemata Kolbe, 1908, p. 29-42. In Beutel, R.G. and Leschen, R.A.B. (eds.), Coleoptera, Beetles. Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). Vol. I, 2 nd edition. Handbook of Zoology Vol. IV, Arthropoda: Insecta. De Gruyter, Berlin, New York.
Hunt, T., Bergsten, J,, Levkanicova, Z., Papadopolou, A., St John, O., Wild, R., Hammond, P.M., Ahrens, D., Balke, M., Caterino, M.S. Gómez-Zurita, J., Ribera, I., Barraclough, T.G., Bocak, L., Vogler, A.P. 2007. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science, 318:1913-1916.
Kirejtshuk, A.G. 1999. Sikhotealinia zhiltzovae (Lafer, 1966) - Recent representative of the Jurassic Coleopterous fauna (Coleoptera, Archostemata, Jurodidae). Proceedings of the Zoological Institute of the Russian Academy of Sciences, 281:21-216.
Kirejtshuk, A.G., Chang, H., Ren, D., and Kun, Sh.Ch. 2010. Family Lasiosynidae n. fam., new paleoendemic Mesozoic family from the infraorder Elateriformia (Coleoptera, Polyphaga). Annales de al Societé Entomologique de France, 46:231-247.
Kirejtshuk, A.G., Nel, A., and Kirejtshuk, P.A. 2016. Taxonomy of the reticulate beetles of the subfamily Cupedinae (Coleoptera: Archostemata) with a review of the historical development. Invertebrate Zoology, 13: 61-190.
Kolbe, H.J. 1908. Mein System der Coleopteren. Zeitschrift für Wissenschaftliche Insektenbiologie, 4:116-123; 5:153-162; 6:219-226; 7:246-251; 8:286-294 10/11:289-400. (in German)
Kukalová-Peck, J. and Lawrence, J.F. 2004. Use of hind wing characters in assessing relationships among coleopteran suborders and major endoneopteran lineages. European Journal of Entomology, 101:95-144.
Lawrence, J.F. 1999. The Australian Ommatidae (Coleoptera: Archostemata): new species, larva and discussion of relationships. Invertebrate Taxonomy, 13: 369-390.
Lawrence, J.F. 2001. A new genus of Valdivian Scirtidae (Coleoptera) with comments on Scirtoidea and the beetle suborders, p. 351-361. In Morimoto, K., Mizuno, K., Hayashi, Y., Ito, T., Ando, N., Tanikado, M., and Shiyake, S. (eds.), Sukunahikona. Special Publication of the Japan Coleopterological Society.
Lawrence, J.F. and Newton, A.F. Jr. 1982. Evolution and classification of beetles. Annual Review of Ecology and Systematics, 12:261-290.
Lawrence, J.F., Ślipiński, S.A., Seago, A.E., Thayer, M.K., Newton, A.F., and Marvaldi, A.E. 2011. Phylogeny of the Coleoptera based on morphological characters of adults and larvae. Annales Zoologici, 61:1-271.
Lawrence, J.F. and Yoshitomi, H. 2007. Nipponocyphon, a new genus of Japanese Scirtidae (Coleoptera) and its phylogenetic significance. Elytra, Tokyo, 35:507-527.
Linnaeus, C. 1758. Systema Naturae per regni tria naturae, secundum classes, ordinaes, genera, species, cum characteribus, differentiis, synonymis, locis. Laurentius Salvius, Stockholm (10th revised ed.). (In Latin)
Martins-Neto, R.G., Gallego, O.F., and Mancuso, A.C. 2006. The Triassic insect fauna from Argentina. Coleoptera from the Los Rastros Formation (Bermejo Basin), La Rioja Province. Ameghiniana, 43:1-26.
Martynov, A.V. 1937. Liassic Insects from Shurab and Kyzyl-Kiya. Trudy Paleontol. Inst. Akad. Nauk SSSR, Moscow-Leningrad. (In Russian)
McKenna, D.D., Wild, A.L., Kanda, K., Bellamy, C.L., Beutel, R.G., Caterino, M.S., Farnum, C.W., Hawks, D.C., Ivie, M.A., Jameson, M.L., Leschen, R.A.B., Marvaldi, A.E., McHugh, J.V., Newton, A.F., Robertson, J.A., Thayer, M.K., Whiting, M.F., Lawrence, J.F., Ślipiński, A., Maddison, D.R., and Farrell, B.D. 2015. The beetle tree of life reveals Coleoptera survived end Permian mass extinction to diversify during the Cretaceous terrestrial revolution. Systematic Entomology, 40:835-880.
Misof, M., Liu, S., Meusemann, K., Peters, R.S., Donath, A., Mayer, C., Frandsen, P.B., Ware, J., Flouri, T., Beutel, R.G., Niehuis, O., Petersen, M., Izquierdo-Carrasco, F., Wappler, T., Rust, J., Aberer, A.J., Aspöck, U., Aspöck, H., Bartel, D., Blanke, A., Berger, S., Böhm, A., Buckley, T., Calcott, B., Chen, J. Friedrich, F., Fukui, M., Fujita, M., Greve, C., Grobe, G., Gu, Sh., Huang, Y., Jermiin, L.S., Kawahara, A.Y., Krogmann, L., Kubiak, M., Lanfear, R., Letsch, H. Li, Y., Li, Zh., Li, J., Lu, H., Machida, R., Mashimo, Y., Kapli, P., McKenna, D.D., Meng, G., Nakagaki, Y., Navarrete-Heredia, J.L., Ott, M., Ou, Y., Pass, G., Podsiadlowski, L., Pohl, H., Reumont, B.M. v., Schütte, K., Sekiya, K., Shimizu, Sh., Slipinski, A., Stamatakis, A., Song, W., Su, X., Szucsich, N.U., Tan, M., Tan, X., Tang, M., Tang, J., Timelthaler, G., Tomizuka, Sh., Trautwein, M., Tong, X., Uchifune, T., Walzl, M.G., Wiegmann, B.M., Wilbrandt, J., Wipfler, B., Wong, T.K.F., Wu, Q., Wu, G., Xie, Y., Yang, Sh., Yang, Q., Yeates, D.K., Yoshizawa, K., Zhang, Q., Zhang, R., Zhang, W., Zhang, Y., Zhao, J., Zhou, Ch., Zhou, L., Ziesmann, T., Zou, Sh., Li, Y., Xu, X., Zhang, Y., Yang, H., Wang, J., Wang, J., Kjer, K.M., Zhou, X. 2014. Phylogenomics resolves the timing and pattern of insect evolution. Science, 346:763-767.
Ponomarenko, A.G. 1963. Paleozoic Beetles Cupedidae of the European Part of the USSR. Paleontological Journal, 1:70-85. (In Russian)
Ponomarenko, A.G. 1968. Jurassic Archostemata beetles of Karatau, p. 118-138. In Rohdendorf, B.B. (ed.), Jurassic insects of Karatau. Nauka, Moscow. (In Russian)
Ponomarenko, A.G. 1969. Historical development of archostematan beetles. Trudy Paleontologicheskogo Instituta, Akademiya Nauk SSSR, Nauka, Moscow. (In Russian)
Ponomarenko A.G. 1977. Composition and ecological characteristics of Mesozoic coleopterans, p. 8-141. In Arnoldy, L.V., Zherikhin, V.V., Nikritin, L.M. and Ponomarenko, A.G. (ed.), Mesozoic Coleoptera. Proceedings of the Academy of Sciences of the USSR. (In Russian).
Ponomarenko, A.G. 1983. Historical development of beetles. Unpublished Habilitation thesis, Paleontological Institute, Moscow, Russian. (In Russian)
Ponomarenko, A.G. 2004. Beetles (Insecta, Coleoptera) of the Late Permian and Early Triassic. Paleontological Journal 38:185-196.
Ponomarenko, A.G. 2008. New Triassic beetles (Coleoptera) from Northern European Russia. Paleontological Journal, 42:600-606.
Ponomarenko, A.G. 2009. The role of arthropods in the development of continental Biota. Paleontological Journal, 43:852-857.
Ponomarenko, A.G., Yan, E.V., Huang, D. 2014. New beetles (Coleoptera) from the terminal Middle Permian of China. Paleontological Journal, 48:191-200.
Rohdendorf, B.B. 1961. Order Coleoptera. Trudy Paleontologicheskogo Instituta, Akademiya Nauk SSSR, 85:393-469.
Scholtz, C.H. and Grebennikov, V.V. 2016. Scarabaeoidea Latrielle, 1802, p. 367-426. In Beutel, R.G. and Leschen, R.A.B. (eds.), Coleoptera, Beetles. Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). Vol. I, 2 nd edition. Handbook of Zoology Vol. IV, Arthropoda: Insecta. De Gruyter, Berlin, New York.
Solier, A.J.J. 1849. Coleópteros. In Gay, C. (ed.), Historia Fisica y Politica de Chile. Zoologia Vol. 4. Privately published, Paris.
Tan, J. and Ren D. 2009. Mesozoic Archostematan Fauna from China. Science Press, Beijing.
Tan, J., Wang, Y., Ren, D., and Yang, X. 2012. New fossil species of ommatids (Coleoptera: Archostemata) from the Middle Mesozoic of China illuminating the phylogeny of Ommatidae. BMC Evolutionary Biology, 12:113.
Tillyard, R.J. and Dunstan, B. 1916. Mesozoic and Tertiary insects of Queenslamd and New South Wales. Queensland Geological Survey Publication, Brisbane, Australia.
Tillyard, R.J. 1924. Upper Permian Coleoptera and a New Order from the Belmont Beds, New South Wales. Proceedings of the Linnean Society of New South Wales, 49:429-435.
Tillyard, R.J. and Dunstan B. 1923. Mesozoic insects of Queensland. Part I. Introduction and Coleoptera. Queensland Geological Survey Publication.
Wallis, J.B. 1928. Revision of the genus Odontaeus, Dej. Canadian Entomologist, 60:119-128; 151-156; 168-176.
Yan, E.V., Wang, B., and Zhang, H. 2013. First record of the beetle family Lasiosynidae (Insecta: Coleoptera) from the Lower Cretaceous of China. Cretaceous Research, 40:43-50.
Yan, E.V., Beutel, R.G., and Ponomarenko, A.G. (2017). †Peltosynidae - a new family from the Middle-Late Triassic of Kirghyzia - its affinities to Polyphaga (Insecta, Coleoptera) and the groundplan of the megadiverse suborder. Journal of Systematic Palaeontology.
dx.doi.org/10.1080/14772019.2017.1313789
