

 An Early Cretaceous Sphenophyllum or a hatchling turtle?
An Early Cretaceous Sphenophyllum or a hatchling turtle?
Article number: 26.3.a50
https://doi.org/10.26879/1306
Copyright Society of Vertebrate Paleontology, December 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 16 May 2023. Acceptance: 25 October 2023.
ABSTRACT
Correctly identifying fossil specimens from parautochtonous deposits where marine and terrestrial organisms co-occur can be challenging due to the abundance of rare and obscure specimens with unclear morphologies. In this study, we reviewed fossils from the Lower Cretaceous La Paja Formation (Ricaurte Alto, Villa de Leyva, Colombia) that were originally described as the plant “Sphenophyllum colombianum” based on an apparent resemblance to the upper Paleozoic genus. We determined that the type specimen corresponds to the carapace of a hatchling turtle. In addition, a second specimen of “S. colombianum” although less well-preserved, also exhibits similar features of a hatchling turtle. The two fossil specimens are significant as they represent the first report of hatchling marine turtles from the Aptian of northwestern South America and provide evidence of the exceptional preservation of the Marine Reptile Lagerstätte of Ricaurte Alto.
Héctor D. Palma-Castro. Departamento de Biología and Grupo de investigación en Paleobiología e Historia Natural, Universidad Nacional de Colombia, Carrera 30 No. 45-03 Bogotá, Colombia and Museo Paleontológico de Villa de Leyva, Universidad Nacional de Colombia, Km 1.5 Vía Arcabuco Villa de Leyva, Colombia. Corresponding author. hdpalmac@unal.edu.co
Diego A. Cómbita-Romero. Departamento de Geociencias and Grupo de investigación en Paleobiología e Historia Natural, Universidad Nacional de Colombia, Carrera 30 No. 45-03 Bogotá, Colombia and Museo Paleontológico de Villa de Leyva, Universidad Nacional de Colombia, Km 1.5 Vía Arcabuco Villa de Leyva, Colombia. dacombitar@unal.edu.co
Edwin-Alberto Cadena. Facultad de Ciencias Naturales and Grupo de Investigación Paleontología Neotropical Tradicional y Molecular (PaleoNeo), Universidad del Rosario, Bogotá, 111221, Colombia and Smithsonian Tropical Research Institute, Box 0843-03092, Balboa, Ancon, Panama, Republic of Panama and Field Museum of Natural History, Chicago, IL, 60605, U.S.A. edwin.cadena@urosario.edu.co
Mónica R. Carvalho. Museum of Paleontology and Department of Earth and Environmental Sciences, University of Michigan, Ann Arbor, Michigan 48109, USA. marvalho@umich.edu
Fabiany Herrera. Earth Sciences, Negaunee Integrative Research Center, Field Museum of Natural History, Chicago, Illinois 60605, USA. fherrera@fieldmuseum.org
Keywords. Aptian; Testudines; Pan-Chelonioidea; South America; Colombia
Final citation: Palma-Castro, Héctor D., Cómbita-Romero, Diego A., Cadena, Edwin-Alberto, Carvalho, Mónica R., and Herrera, Fabiany. 2023. An Early Cretaceous Sphenophyllum or a hatchling turtle? Palaeontologia Electronica, 26(3):a50.
https://doi.org/10.26879/1306
palaeo-electronica.org/content/2023/4017-fossil-plant-or-turtle
Copyright: December 2023 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
According to the most recent list published by the International Union of Geological Sciences (IUGS), only two Geological Heritage Sites have been recognized in Colombia (IUGS, 2022): the Nevado del Ruiz Quaternary Volcanic Complex and the Marine Reptile Lagerstätte from the Lower Cretaceous of the Ricaurte Alto. The Ricaurte Alto is a coastal and shallow marine sequence comprising approximately 1.9 km of sedimentary rocks from the Upper Valanginian to the Albian. It is divided into four formations (Ritoque Formation, La Paja Formation, San Gil Inferior Formation, and San Gil Superior Formation) as described by Etayo-Serna (1968).
Within the Ricaurte Alto sequence, La Paja Formation has yielded an enormous quantity and diversity of exceptionally preserved Hauterivian to Aptian fossils (~132 to ~113 Ma; Etayo-Serna, 1968). These fossils include a large number of marine reptiles, such as plesiosaurs (Welles, 1962; Páramo-Fonseca et al., 2019), pliosaurs (Páramo-Fonseca et al., 2016, 2018; Gómez-Pérez and Noè, 2017; Noè and Gómez-Pérez, 2022;), ichthyosaurs (Páramo-Fonseca, 1997; Maxwell et al., 2016; Páramo-Fonseca et al., 2020; Cortés et al., 2021), turtles (Cadena, 2015; Cadena and Parham, 2015; Cadena et al., 2019), crocodylomorphs (Cortés et al., 2019), and dinosaurs (Carballido et al., 2015; Cortés et al., 2023). Marine invertebrates are also widespread in the sedimentary sequence, particularly ammonoids (Etayo-Serna, 1968, 1979; Patarroyo, 2000, 2020) and orithopsid crabs (Luque et al., 2020).
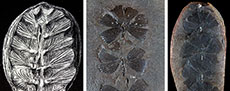
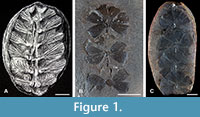 Plant remains are sporadically found in calcareous concretions and shale beds of La Paja Formation. These plants likely reflect elements of the coastal vegetation that were capable of withstanding transportation. Over 50 plant taxa have been described (Huertas, 1967, 1970a, 1976, 2003; van Waveren et al., 2002; Moreno et al., 2007), however, most of their descriptions and affinities require a systematic review (Palma, 2020). In his last work, Huertas (2003) described abundant cast-mold and permineralized remains of stems, branches, leaves, and reproductive structures of gymnosperm affinity. Most plants appear to be related to Araucariaceae, Cupressaceae, and Bennettitales (van Waveren et al., 2002). Among these plants, a presumed sphenophyte species, “Sphenophyllum colombianum” (Huertas, 2003) (Figure 1A), was also described from La Paja Formation, far extending the temporal and stratigraphic range known for this Paleozoic genus. Sphenophyllum is an extinct plant found worldwide, ranging from the Late Devonian to the Permian and exhibiting a large morphological diversity during the Carboniferous (Taylor et al., 2009; de Felice et al., 2019; Huang et al., 2022; Rischbieter et al., 2022). Well-identified leaves of Sphenophyllum (Figure 1B-C) are borne in whorls and typically show wedge-shaped forms with veins radiating from the leaf base (Taylor et al., 2009). The unexpected discovery of a species of Sphenophyllum in the La Paja Formation represents a conundrum: either this Paleozoic genus extended into the Early Cretaceous or the affinity of “S. colombianum” is incorrect.
Plant remains are sporadically found in calcareous concretions and shale beds of La Paja Formation. These plants likely reflect elements of the coastal vegetation that were capable of withstanding transportation. Over 50 plant taxa have been described (Huertas, 1967, 1970a, 1976, 2003; van Waveren et al., 2002; Moreno et al., 2007), however, most of their descriptions and affinities require a systematic review (Palma, 2020). In his last work, Huertas (2003) described abundant cast-mold and permineralized remains of stems, branches, leaves, and reproductive structures of gymnosperm affinity. Most plants appear to be related to Araucariaceae, Cupressaceae, and Bennettitales (van Waveren et al., 2002). Among these plants, a presumed sphenophyte species, “Sphenophyllum colombianum” (Huertas, 2003) (Figure 1A), was also described from La Paja Formation, far extending the temporal and stratigraphic range known for this Paleozoic genus. Sphenophyllum is an extinct plant found worldwide, ranging from the Late Devonian to the Permian and exhibiting a large morphological diversity during the Carboniferous (Taylor et al., 2009; de Felice et al., 2019; Huang et al., 2022; Rischbieter et al., 2022). Well-identified leaves of Sphenophyllum (Figure 1B-C) are borne in whorls and typically show wedge-shaped forms with veins radiating from the leaf base (Taylor et al., 2009). The unexpected discovery of a species of Sphenophyllum in the La Paja Formation represents a conundrum: either this Paleozoic genus extended into the Early Cretaceous or the affinity of “S. colombianum” is incorrect.
Here, we re-examined the material described as “Sphenophyllum colombianum” by Huertas (2003) and additional material found in the Huertas’ collection related to the same species. Our reevaluation shows that the specimens described as “S. colombianum” are unequivocal carapaces of hatchling turtles.
MATERIAL AND METHODS
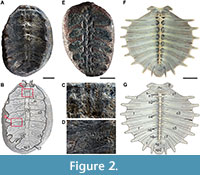 The “type” specimen of “Sphenophyllum colombianum” described by Huertas is an open and slightly eroded calcareous concretion (Figure 2A-D). A second specimen with similar characteristics, labeled as a “Cotypus” was also analyzed (Figure 2E). Both specimens are housed in the paleontological collections of the Department of Geosciences at the Universidad Nacional de Colombia (UNDG). The “type” specimen (MONQ-602) was obtained from Monquirá, near the “Estación del Kronosaurio” in Villa de Leyva (~5°38’13” N, ~73°33’33” W), while the second specimen (LLC-65) was collected from La Catalina Hill, located on the border between the towns of Villa de Leyva and Sutamarchán (~5°38’2” N, ~73°34’40” W).
The “type” specimen of “Sphenophyllum colombianum” described by Huertas is an open and slightly eroded calcareous concretion (Figure 2A-D). A second specimen with similar characteristics, labeled as a “Cotypus” was also analyzed (Figure 2E). Both specimens are housed in the paleontological collections of the Department of Geosciences at the Universidad Nacional de Colombia (UNDG). The “type” specimen (MONQ-602) was obtained from Monquirá, near the “Estación del Kronosaurio” in Villa de Leyva (~5°38’13” N, ~73°33’33” W), while the second specimen (LLC-65) was collected from La Catalina Hill, located on the border between the towns of Villa de Leyva and Sutamarchán (~5°38’2” N, ~73°34’40” W).
The fossils were photographed using a Canon EOS R camera with MP-E 65mm f/2.8 1-5x and TS-E 50mm f/2.8L Macro lenses attached to a StackShot system at the Universidad Nacional de Colombia, Bogota. The digital images were merged using Helicon Focus software.
The fossils were compared with specimens of extant marine turtles, including Eretmochelys imbricata MTKD-8295 (Senckenberg Museum, Dresden), Caretta caretta AMNH-129869 (American Museum of Natural History, New York), and Lepidochelys olivacea QM-J85545 (Queensland Museum, Brisbane). Specimens of Sphenophyllum emarginatum (PP-16865 and PP-58015) from the Mazon Creek flora, Carbondale Formation (Pennsylvanian) were photographed at the Field Museum (Chicago, USA) for comparison.
RESULTS
Upon close examination, the specimens of “Sphenophyllum colombianum” do not represent a fossil plant but rather the remains of carapaces of hatchling marine turtles. The specimen MONQ-602 (Figure 2A-D) corresponds to a partially preserved, small carapace (~6.1 cm long) in ventral view, missing all the peripherals, the anteromedial portion of the nuchal, the suprapygal 2, and the pygal. It preserves at least seven neural bones, seven costal bones, suprapygal 1, and the most posterior portion of the nuchal (Figure 2A-B). The sutural contact between these bones is serrated (Figure 2C-D), however, many of the neural and costal sutural contacts can only be inferred, especially those involving the posterior neurals after neural 4 (Figure 2B). The rib head that articulates with the missing thoracic vertebrae are also visible and well defined. The bone surface is characterized by an irregular pitted texture and radial bone growth patterns, which Huertas (2003) described as leaf venation (Figure 2B, D). MONQ-602 also shows part of the fontanelles between lateral portion of costals and the missing peripherals, and an oval-shaped space between the nuchal and the left costal 1 that resembles the post-nuchal fontanel (Figure 2B).
The specimen LLC-65 (Figure 2E) corresponds to the impression of a poorly ossified carapace (~5 cm long) in ventral view, which does not show a clear bone texture or growth pattern. However, the rough impressions of the neurals, costals (rounded due to the poor preservation), and the scars for the insertion of thoracic vertebrae into the neurals are discernable.
DISCUSSION
Misidentified Fossil Plants from Colombia
The re-examination of “Sphenophyllum colombianum” indicates that this species is invalid as it was based on misinterpreted skeletal remains of marine turtles. The hypothesized occurrence of Sphenophyllum in the Early Cretaceous of Colombia would have implied that the genus survived beyond the Permian in northwest Gondwana. However, the new evidence presented here cannot uphold this interpretation. Other incorrectly identified fossil plants from Villa de Leyva include the cocoa plant “Theobroma fossilium” Berry (1929), which is actually the jaw fragment of a marine reptile (Brown, 1946; Huertas, 1971). Seeds of “Musa enseteformis” Berry (1925) from Bogotá, later transferred to Ensete enseteformis (Berry) Jain (1965), turned out to be modern seeds of E. ventricosum (Manchester and Kress, 1993). Similarly, a presumed Cretaceous fruit of Musaceae (Huertas and van der Hammen, 1953; Huertas, 1970b) was, in fact, a banana-shaped sedimentary concretion found in Cundinamarca (Manchester and Kress, 1993). All these misidentified fossils that are historically attached to the paleobotanical literature support the re-examination of other critical fossils that Huertas (2003) described from the La Paja Formation.
Age of the Fossils
The age of these fossils is re-interpreted based on previous and new geological data. Huertas (2003) considered the fossils as Barremian in age. However, this is highly unlikely because the ages near the “Estación del Kronosaurio” and the outcrop from the La Catalina Hill are late Aptian or Aptian based on ammonoids (Etayo-Serna, 1968; Acosta et al., 1979; Forero and Sarmiento L., 1985; Noè and Gómez-Pérez, 2022;, Benavides-Cabra et al., 2023). Therefore, MONQ-602 and LLC-65 turtle specimens are now interpreted as late Aptian or Aptian in age.
Taxonomic Identification of the Fossils
MONQ-602 and LLC-65 specimens from La Paja Formation resemble hatchlings and juveniles of both extant and extinct marine turtles in many aspects, including a relatively poor ossification, highly serrated sutural contacts between bones, and the presence of fontanelles (Reisz and Head, 2008). We can exclude the possibility that they belong to pleurodirans (side-necked turtles) due to the lack of evidence of an iliac scar on the posterior costal bones, which suggests that the illium was not fused to the carapace. The iliac scar is present in hatchling/juvenile pleurodirans, like Puentemys mushaisaensis from the Paleocene of Colombia (Cadena et al., 2012). In addition, the continuous series of neurals reaching the suprapygal 1 exclude them from being pan-pelomedusoids.
Overall, the skeletal and morphological characteristics preserved in both fossil specimens match those exhibited by the carapace’s ventral surface of extant marine turtles, such as those seen in Lepidochelys olivacea (Figure 2F-G). However, MONQ-602 shows fewer neurals and costals than expected for a turtle of this size, seven instead of the expected eight. It is possible that a very small neural 8 and narrow costals 8 with weak sutural contact may exist in the specimen but are not discernible due to poor preservation of this posterior region. Another hypothesis is that the structure that we interpret as the nuchal, could correspond to the posterior portion of neural 1. However, this hypothesis seems to be invalid, as what we attribute as neural 1 is restricted to the first pair of costals, as seen in other pan-chelonioids and the Lepidochelys olivacea specimen shown in Figure 2G.
Giving the co-occurrence of the two specimens re-examined here and other mature turtles in La Paja Formation, it is plausible to suggest that the specimens correspond to hatchlings of the protostegid Desmatochelys padillai Cadena and Parham (2015). D. padillai is a marine fossil turtle abundant in La Paja Formation, and nearly complete specimens of adults have been found (Cadena and Parham, 2015). However, in the absence of fully preserved skeletons including skulls, we refrain from establishing a detailed systematic paleontology, and for now, the fossils should be considered as Pan-Chelonioidea indet. turtles.
Until now, only a few Cretaceous marine pan-chelonioid hatchlings and juvenile specimens have been described from the early Campanian of USA (Zangerl, 1953; Matzke, 2007; Gentry, 2018), and late Middle Cenomanian of Lebanon (Tong et al., 2006). The MONQ-602 specimen shows intercostal contacts, distinguishing it from other protostegid hatchlings such as Rhinochelys nammourensis (Tong et al., 2006). However, other hatchling turtles outside Pan-Chelonioidea also lack intercostal contacts, as observed in the pelomedusoid Araripemys barretoi (Oliveira and Kellner, 2017). This indicates that this characteristic cannot be used as diagnostic or exclusive of hatchling protostegids.
Ontogenetic Stage Determination
Aside from species-specific ontogenetic distinctions, two common co-occurring characteristics can help in identifying hatchling turtles in the fossil record: (i) skeletal immaturity, characterized by reduced ossification and the presence of large fontanelles, and (ii) small size (Lindgren et al., 2017). By examining the correlation between body size and age in extant turtles (Chaloupka and Zug, 1997; Zug et al., 2002), we can hypothesize estimates of the ontogenetic stage of the MONQ-602 and LLC-65 specimens.
Extant marine turtle hatchlings typically grow to have a curved carapace length (CCL) of 4 to 6 cm within their first year. On average, after this initial year, hatchlings transition into the juvenile stage in which their lengths increase beyond 17 cm. Adulthood is reached at around 14 years of age, when straight carapace length (SCL) measurement exceeds 40 cm (Snover et al., 2007; Avens et al., 2009; Avens et al., 2012; Avens et al., 2021; Turner-Tomaszewicz et al., 2022; Baldi et al., 2023) (Table 1). The specimen MONQ-602 is ~6.1 cm long and LLC-65 is ~5 cm long. Based on these measurements and the growth patterns observed on living turtles, the La Paja specimens were likely in the hatchling/post-hatchling stage, implying that they were probably between 0-1 years old at the moment of death. However, it is important to point out that the size of marine turtles is affected by various factors, and not all living marine turtles exhibit the same growth patterns or reach the same sizes in adulthood. For instance, Dermochelys corieracea grows faster and reaches a larger size than all other living marine turtles. Similarly, protostegid turtles show a consistent evolutionary trend towards larger size. Recently, Wilson (2023) demonstrated that the osteohistology of Protostega gigas closely resembles that of D. corieracea, suggesting that both species share a rapid early ontogenetic growth, while Desmatochelys lowi (an Early Cretaceous protostegid) (Elliott et al., 1997) lacks the same quick growth patterns, leading to the conclusion that this strategy likely evolved within the protostegid lineage and is not an ancestral trait of the clade. The presence of Valanginian large protostegid shells and limb bones provides evidence that this turtle lineage rapidly evolved large body size during the Early Cretaceous (Cadena and Combita-Romero, 2023).
In addition to their size, the skeletal immaturity observed in MONQ-602 and LLC-65 specimens is discernible. The ontogenetic stage is comparable to that of extant turtle hatchlings, such as four weeks old Chelonia mydas, as documented by Zug et al. (2002). The presence of large fontanelles and very low carapace thickness due to poor dermal ossification supports the interpretation that these fossil specimens represent hatchlings. The specimen MONQ-602 has strong and well-developed intercostal sutural contacts, different from other smaller fossil hatchling carapaces, like the protostegid Rhinochelys nammourensis (Tong et al., 2006) and the pelomedusoid Araripemys barretoi (Oliveira and Kellner, 2017). This suggests that MONQ-602 probably died in a post-hatchling stage when its carapace was slightly more developed than that of specimen LLC-65.
Taphonomic Insights
Although La Paja Formation hatchling turtles slightly differ in size (specimen MONQ-602 is somewhat larger than LLC-65), both fossils exhibit similar modes of preservation. Most of the outer bones of the carapace, including the anteromedial portion of the nuchal, all peripherals, suprapygal 2, and pygal are missing. This suggests that these elements, along with cranial and limb bones, were lost prior to burial and subsequent fossil diagenesis through concretion formation. In addition, both concretions show no evidence of marginal breaks, only moderate abrasion, which excludes the possibility that the peripheral bone elements were lost during a later erosional event. Furthermore, there is no evidence of anatomical displacement between the preserved bone elements (neurals, costals, and suprapygal 1) in both specimens, indicating minimal disarticulation. Considering studies of extant turtle disarticulation (Brand et al, 2003), the preservation of the Paja hatchlings suggests that the burial was likely rapid, occurring a few weeks after death.
CONCLUSIONS
The fossils described originally as “Sphenophyllum colombianum” by Huertas, from the La Paja Formation in Colombia, have now been identified as hatchling marine turtles belonging to Pan-Chelonioidea indet. This discovery represents the first report of hatchlings turtle carapaces from northwestern South America. MONQ-602 specimen is a new example of the outstanding record of turtles from the Ricaurte Alto Lagerstätte that now includes eggs, hatchings, and completely articulated adults. Further preparation and potential micro-CT scanning of specimen MONQ-602 will likely allow the identification of the dorsal surface of the carapace and explore if other bone remains were also preserved in the concretion. This work aims to promote future collaborative work between different paleontological disciplines, such as paleobotany and vertebrate paleontology, to prevent taxonomical misidentifications, especially when dealing with fossils that are partially complete or pose specific identification challenges.
ACKNOWLEDGMENTS
We thank the Paleontological collection of the Universidad Nacional de Colombia for allowing us the examination of these fossils. None of the authors have any type of conflict of interest. This project was supported by the National Geographic Society, grant (EC-96755R-22) Discovering Early Cretaceous Floras from Northern South America to FH and the Field Museum Visiting Scholarship (Negaunee Integrative Research Center) to HDP-C. We thank Dr. S. Evers and an anonymous reviewer for their valuable comments.
REFERENCES
Acosta, C.E., Huertas, G., and Ruiz, P.M. 1979. Noticia preliminar sobre el hallazgo de un presunto Kronosaurus (Reptilla: Dolichrorhynchopidae) en el Aptiano superior de Villa de Leiva Colombia. Lozanía (Acta Zoologica Colombiana), 28:1–7.
Avens, L., Taylor, J.C., Goshe, L.R., Jones, T.T., and Hastings, M. 2009. Use of skeletochronological analysis to estimate the age of leatherback sea turtles Dermochelys coriacea in the western North Atlantic. Endangered Species Research, 8:165–177.
https://doi.org/10.3354/esr00202
Avens, L., Goshe, L.R., Harms, C.A., Anderson, E.T., Hall, A.G., Cluse, W.M., Godfrey, M.H., Braun-McNeill, J., Stacy, B., Bailey, R., and Lamont, M.M. 2012. Population characteristics, age structure, and growth dynamics of neritic juvenile green turtles in the northeastern Gulf of Mexico. Marine Ecology Progress Series, 458:213–229.
https://doi.org/10.3354/meps09720
Avens, L., Ramirez, M.D., Goshe, L.R., Clark, J.M., Meylan, A.B., Teas, W., and Howell, L. 2021. Hawksbill sea turtle life-stage durations, somatic growth patterns, and age at maturation. Endangered Species Research, 45:127–145.
https://doi.org/10.3354/esr01123
Baldi, G., Furii, G., Del Vecchio, M., Salvemini, P., Vallini, C., Angelini, V., Pari, S., Moraes, K.L., Margaritoulis, D., Çurri, A., Lazar, B., and Casale, P. 2023. Growth rates and age at maturity of Mediterranean loggerhead sea turtles estimated from a single-population foraging ground. Marine Biology, 170(4): 36.
https://doi.org/10.1007/s00227-023-04189-1
Benavides-Cabra, C.D., Páramo-Fonseca, M.E., Palma-Castro, H.D., Narváez-Rincón, J.A., and Ramos-Clavijo, M.P. 2023. Stratigraphic distribution of marine vertebrates of the Arcillolitas Abigarradas member (Paja Formation) from the Villa de Leiva - Sáchica - Sutamarchán region, Boyacá, Colombia. Earth Sciences Research Journal, 27 (3), 211-226.
https://doi.org/10.15446/esrj.v27n2.108292
Berry, E.W. 1925. A banana in the Tertiary of Colombia. American Journal of Science, 5(60):530–537.
Berry, E.W. 1929. Tertiary fossil plants from Colombia, South America. Proceedings of the United States National Museum, 75(2795):1–12.
Brand, L.R., Hussey, M., and Taylor, J. 2003. Taphonomy of freshwater turtles: decay and disarticulation in controlled experiments. Journal of Taphonomy, 1(4):233–245.
Brown, R.W. 1946. Alterations in some fossil and living floras. Journal of the Washington Academy of Sciences, 36(10):344–355.
Cadena, E.A. 2015. The first South American sandownid turtle from the Lower Cretaceous of Colombia. PeerJ, 3:e1431.
https://doi.org/10.7717/peerj.1431
Cadena, E.A. and Combita-Romero, D.A. 2023. The onset of large size in Cretaceous marine turtles (Protostegidae) evidenced by new fossil remains from the Valanginian of Colombia. Zoological Journal of the Linnean Society.
https://doi.org/10.1093/zoolinnean/zlad053
Cadena, E.A. and Parham, J.F. 2015. Oldest known marine turtle? A new protostegid from the Lower Cretaceous of Colombia. PaleoBios, 32:1–42.
https://doi.org/10.11436/mssj.15.250
Cadena, E.A., Bloch, J.I., and Jaramillo, C.A. 2012. New bothremydid turtle (Testudines, Pleurodira) from the Paleocene of northeastern Colombia. Journal of Paleontology, 86(4):688–698.
https://doi.org/10.1666/11-128R1.1
Cadena, E.A., Parra-Ruge, M.L., Parra-Ruge, J. de D., and Padilla-Bernal, S. 2019. A gravid fossil turtle from the Early Cretaceous reveals a different egg development strategy to that of extant marine turtles. Palaeontology, 62(4):533–545.
https://doi.org/10.1111/pala.12413
Carballido, J.L., Pol, D., Parra Ruge, M.L., Padilla Bernal, S., Paramo-Fonseca, M.E., and Etayo-Serna, F. 2015. A new Early Cretaceous brachiosaurid (Dinosauria, Neosauropoda) from northwestern Gondwana (Villa de Leiva, Colombia). Journal of Vertebrate Paleontology, 35(5):e980505.
https://doi.org/10.1080/02724634.2015.980505
Cortés, D., Larsson, H.C.E., Maxwell, E.E., Parra Ruge, M.L., Patarroyo, P., and Wilson, J.A. 2019. An Early Cretaceous Teleosauroid (Crocodylomorpha: Thalattosuchia) from Colombia. Ameghiniana, 56(5):365–379.
https://doi.org/10.5710/AMGH.26.09.2019.3269
Cortés, D., Maxwell, E.E., and Larsson, H.C.E. 2021. Re-appearance of hypercarnivore ichthyosaurs in the Cretaceous with differentiated dentition: revision of ‘ Platypterygius ’ sachicarum (Reptilia: Ichthyosauria, Ophthalmosauridae) from Colombia. Journal of Systematic Palaeontology, 19(14):969–1002.
https://doi.org/10.1080/14772019.2021.1989507
Cortés, D., Parra-Ruge, M., Parra-Ruge, J., Deers-Potvin, A., Smith, A., Hoai-Nam, B., and Larsson, H. 2023. Theropod dinosaur footprints from the early cretaceous of Colombia: walking dynamics and paleoenvironmental implications. Facies, 9:1–17.
Chaloupka, M. and Zug, G.R. 1997. A polyphasic growth function for the endangered Kemp’s ridley sea turtle, Lepidochelys kempii. Fishery Bulletin, 95:849–856.
de Felice, H.T., Decombeix, A.-L., and Galtier, J. 2019. Anatomy, affinities, and evolutionary implications of new silicified stems of Sphenophyllum Brongniart, 1828 from the early Carboniferous (Mississippian) of France and Germany. Geodiversitas, 41(1):587.
https://doi.org/10.5252/geodiversitas2019v41a14
Elliott, D. K., Irby, G. V., and Hutchison, J. H. 1997. Desmatochelys lowi, a marine turtle from the Upper Cretaceous. p. 243—258. In Callaway, J. and Nicholls, L. (eds.), Ancient marine reptiles, Academic Press.
https://doi.org/10.1016/B978-012155210-7/50014-4
Etayo-Serna, F. 1968. Sinopsis estratigráfica de la región de Villa de Leiva y zonas próximas. Boletín de Geología, 21:19–32.
Etayo-Serna, F. 1979. Zonation of the Cretaceous of central Colombia by Ammonites. Publicaciones Geológicas Especiales, 2, Ingeominas, Bogotá, Colombia.
Forero, H. and Sarmiento L. 1985. Las facies evaporíticas de la Formación Paja en la región de Villa de Leyva, XVII, p. 16. In Etayo-Serna, F. and Laverde, F. (eds.), Proyecto Cretácico, Publicaciones Geológicas Especiales, 16, Ingeominas, Bogotá, Colombia.
Gentry, A.D. 2018. Prionochelys matutina Zangerl, 1953 (Testudines: Pan-Cheloniidae) from the Late Cretaceous of the United States and the evolution of epithecal ossifications in marine turtles. PeerJ, 6:e5876.
https://doi.org/10.7717/peerj.5876
Gómez-Pérez, M. and Noè, L.F. 2017. Cranial anatomy of a new pliosaurid Acostasaurus pavachoquensis from the Lower Cretaceous of Colombia, South America. Palaeontographica, Abteilung A, 310 (1-2):5–42.
https://doi.org/10.1127/pala/2017/0068
Hirayama, R. 1998. Oldest known sea turtle. Nature, 392 (6677):705–708.
Huang, P., Liu, L., Liu, L., Wang, J.S., and Xue, J.Z. 2022. Sphenophyllum Brongniart (Sphenopsida) from the Upper Devonian of South China. Palaeoworld, 31(3):402–418.
https://doi.org/10.1016/J.PALWOR.2021.09.007
Huertas, G. 1967. Sertum florulae fossilis Villae de Leivae. Caldasia, 9 (46):59–75.
Huertas, G. 1970a. Sertum florulae fossilis Villae de Leiva II. Caldasia, 10 (50):595–602.
Huertas, G. 1970b. Disquisición Paleobotánica. Mutisia, 23:21–28.
Huertas, G. 1971. Theobroma verum Huertas sp. nov. Mutisia, 34:10–15.
Huertas, G. 1976. Sertum florulae fossilis Villae de Leiva. Caldasia, 11(54):17–23.
Huertas, G. 2003. Flora Fósil de Villa de Leyva y sus alrededores. Camargo Editores, Chía, Colombia.
Huertas, G. and van der Hammen, T. 1953. Un posible banano (Musa) fosil del Cretáceo de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Fisicas y Naturales, 9:115–117.
IUGS. 2022. Designations. IUGS Geological Heritage Sites. Visited September 16 2022.
https://iugs-geoheritage.org/designations/
Jain, R.K. 1965. Studies in Musaceae: III. Fossil records of Musaceae and the origin of bananas. Proceedings/Indian Academy of Sciences, 61 (3):170–179.
Lindgren, J., Kuriyama, T., Madsen, H., Sjövall, P., Zheng, W., Uvdal, P., Engdahl, A., Moyer, A.E., Gren, J.A., Kamezaki, N., Ueno, S., and Schweitzer, M.H. 2017. Biochemistry and adaptive colouration of an exceptionally preserved juvenile fossil sea turtle. Scientific Reports, 7(1):1–13.
https://doi.org/10.1038/s41598-017-13187-5
Luque, J., Cortés, D., Rodriguez-Abaunza, A., Cárdenas, D., and Parra, J de D. 2020. Orithopsid crabs from the Lower Cretaceous Paja Formation in Boyacá (Colombia), and the earliest record of parasitic isopod traces in Raninoida. Cretaceous Research, 116:104602.
https://doi.org/10.1016/j.cretres.2020.104602
Manchester, S.R., and Kress, W.J. 1993. Fossil bananas (Musaceae): Ensete oregonense sp. nov. from the Eocene of western North America and its phytogeographic significance. American Journal of Botany, 80(11):1264–1272.
Matzke, A.T. 2007. An almost complete juvenile specimen of the cheloniid turtle Ctenochelys stenoporus (Hay, 1905) from the Upper Cretaceous Niobrara Formation of Kansas, USA. Palaeontology, 50(3):669–691.
https://doi.org/10.1111/j.1475-4983.2007.00650.x
Maxwell, E.E., Dick, D., Padilla, S., and Parra, M.L. 2016. A new ophthalmosaurid ichthyosaur from the Early Cretaceous of Colombia. Papers in Palaeontology, 2(1):59–70.
https://doi.org/10.1002/spp2.1030
Moreno, M., Cruz, A. de J.G., and González, H.C. 2007. Frenelopsis y Pseudofrenelopsis (Coniferales: Cheirolepidiaceae) en el Cretácico Temprano de Colombia. Boletín de Geología, 29(2):13–19.
Noè, L.F., and Gómez-Pérez, M. 2022. Giant pliosaurids (Sauropterygia; Plesiosauria) from the Lower Cretaceous peri-Gondwanan seas of Colombia and Australia. Cretaceous Research, 132:105122.
https://doi.org/10.1016/J.CRETRES.2021.105122
Oliveira, G.R. and Kellner, A.W. 2017. Rare hatchling specimens of Araripemys Price, 1973 (Testudines, Pelomedusoides, Araripemydidae) from the Crato Formation, Araripe Basin. Journal of South American Earth Sciences, 79:137–142.
Palma Castro, H.D. 2020. Primer reporte de una conífera en el Aptiano de Vélez, Santander, consideraciones tafonómicas, Cretácico Inferior de Colombia. Unpublished undergraduate thesis project. Universidad Nacional de Colombia, Bogotá, Colombia.
Páramo-Fonseca, M.E. 1997. Platypterigius sachicarum (Reptilia, ichthyosauria) nueva especia del cretácico de Colombia. Revista Ingeominas, 6:1–12.
Páramo-Fonseca, M.E., Benavides-Cabra, C.D., and Gutiérrez, I.E. 2018. A new large pliosaurid from the Barremian (Lower cretaceous) of Sáchica, Boyacá, Colombia. Earth Sciences Research Journal, 22(4):223–238.
https://doi.org/10.15446/esrj.v22n4.69916
Páramo-Fonseca, M.E., García-Guerrero, J., Benavides-Cabra, C.D., Padilla-Bernal, C., and Castañeda-Gómez, J.A. 2020. A benchmark specimen of Muiscasaurus catheti from the upper Aptian of Villa de Leiva, Colombia: New anatomical features and phylogenetic implications. Cretaceous Research, 119:104685.
https://doi.org/10.1016/j.cretres.2020.104685
Páramo-Fonseca, M.E., Gómez-Pérez, M., Noè, L.F., and Etayo-Serna, F. 2016. Stenorhynchosaurus munozi, gen. et sp. nov. a new pliosaurid from the Upper Barremian (Lower Cretaceous) of Villa de Leiva, Colombia, South America. Revista de la Academia Colombiana de Ciencias Exactas, Fisicas y Naturales. 40(154):84–103.
Páramo-Fonseca, M.E., O’Gorman, J.P., Gasparini, Z., Padilla, S., and Parra-Ruge, M.L. 2019. A new late Aptian elasmosaurid from the Paja Formation, Villa de Leiva, Colombia. Cretaceous Research, 99:30–40.
https://doi.org/10.1016/J.CRETRES.2019.02.010
Patarroyo, P. 2000. Distribución de amonitas del Barremiano de la Formación Paja en el sector de Villa de Leyva (Boyacá, Colombia). Geología Colombiana, 25:149–162.
Patarroyo, P. 2020. Barremian Deposits of Colombia: A Special Emphasis on Marine Succession, p. 1–37. In Gómez, J. and Pinilla-Pachon, A.O. (eds.), The Geology of Colombia, Volume 2 Mesozoic. Servicio Geológico Colombiano, Publicaciones Geológicas Especiales, Bogotá.
https://doi.org/10.32685/pub.esp.36.2019.12
Reisz, R.R. and Head, J.J. 2008. Turtle origins out to sea. Nature, 456: 450–451.
https://doi.org/10.1038/456450a
Rischbieter, M., Neregato, R., Iannuzzi, R., di Pasquo, M.M., Alvarenga, R., and Freitas, J. 2022. A new flora from the Rio Bonito Formation (late Asselian) and its implications for the biostratigraphy of the southern Paraná Basin, Brazil. Journal of South American Earth Sciences, 119:104010.
https://doi.org/10.1016/J.JSAMES.2022.104010
Snover, M.L., Hohn, A.A., Crowder, L.B., and Heppell, S.S. 2007. Age and growth in Kemp’s ridley sea turtles: evidence from mark-recapture and skeletochronology, p. 89–106. In Plotkin, P. T. (ed.), Biology and conservation of ridley sea turtles, Johns Hopkins University Press.
Taylor, E.L., Taylor, T.N., and Krings, M. 2009. Paleobotany: the biology and evolution of fossil plants. Academic Press.
Tong, H., Hirayama, R., Makhoul, E., and Escuillié, F. 2006. Rhinochelys (Chelonioidea: Protostegidae) from the Late Cretaceous (Cenomanian) of Nammoura, Lebanon. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano, 147:113–38.
Turner-Tomaszewicz, C.N., Avens, L., Seminoff, J.A., Limpus, C.J., FitzSimmons, N.N., Guinea, M.L., Pendoley, K.L., Whittock, P.A., Vitenbergs, A., Whiting, S.D., and Tucker, A.D. 2022. Age-specific growth and maturity estimates for the flatback sea turtle (Natator depressus) by skeletochronology. PLoS ONE, 17(7):e0271048.
https://doi.org/10.1371/journal.pone.0271048
van Waveren, I.M., van Konijnenburg-Van Cittert, J.H.A., van den Burgh, J., and Dilcher, D. L. 2002. Macrofloral remains from the Lower Cretaceous of the Leiva region (Colombia). Scripta Geologica, 123:1–39.
Welles, S.P. 1962. A new species of elasmosaur from the Aptian of Colombia and a review of the Cretaceous plesiosaurs. University of California Publications in the Geological Sciences, 44: 1–96.
https://doi.org/10.14825/KASEKI.85.0_69
Zangerl, R. 1953. The vertebrate fauna of the Selma Formation of Alabama. Part 3. The turtles of the family Protostegidae. Part 4. The turtles of the family Toxochelyidae. Fieldiana, Geology, Memoirs, 3:61–277.
Zug, G.R., Balazs, G.H., and Parker, D.M. 2002. Age and growth of Hawaiian green seaturtles (Chelonia mydas): an analysis based on skeletochronology. Fishery Bulletin, 100:117–127.
