

 The first record of Lower Cretaceous otoliths from the Kimigahama Formation (Barremian) of the Choshi Group, Chiba Prefecture, Japan
The first record of Lower Cretaceous otoliths from the Kimigahama Formation (Barremian) of the Choshi Group, Chiba Prefecture, Japan
Article number: 27.1.a20
https://doi.org/10.26879/1318
Copyright Society of Vertebrate Paleontology, April 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 9 July 2023. Acceptance: 25 February 2024.
ABSTRACT
The Lower Cretaceous otolith assemblage from the Kimigahama Formation (Barremian) of the Choshi Group, Chiba Prefecture, Japan, is described. The Kimigahama Formation consists of bioturbated silty sandstone, sandy siltstone, and hummocky and swaley cross-stratified sandstone, representing an offshore to shoreface depositional environment. A total of five otolith types were identified, including undetermined Teleostei, Pterothrissinae, Elopiformes, Argentinidae, and Ichthyotringidae. Among them, Ichthyotringidae from Kimigahama Formation is the oldest Ichthyotringoidei. Based on the shallow marine setting of the Kimigahama Formation, the Barremian Pterothrissinae and Argentinidae in East Asia inhabited shallow marine environments, whereas the habitat of extant species is the deep sea. Furthermore, our new data on Pterothrissinae, Argentinidae, Ichthyotringoidei, and Elopiformes from Barremian strata in the northwestern Pacific indicate that these taxa exhibited a cosmopolitan distribution in the Pacific, Tethys, and Atlantic seas during the middle Early Cretaceous.
Shinya Miyata. Oishi Fossil Gallery of Mizuta Memorial Museum Josai University Educational Corporation, 2-3-20 Hirakawa-cho, Chiyoda-ku, Tokyo 102-0093, Japan and Faculty of Science, Josai University 1-1 Keyakidai Sakado, Saitama 350-0295, Japan.
fossil.paleoichthyology777@gmail.com
Shinji Isaji. Natural History Museum and Institute, Chiba, 955-2 Aoba-cho, Chuo-ku, Chiba 260-8682, Japan. isaji@chiba-muse.or.jp
Kenji Kashiwagi.Department of Natural and Environmental Sciences, School of Science,University of Toyama, 3190 Gofuku, Toyama 930-855, Japan. kasiwagi@sci.u-toyama.ac.jp
Hidehiko Asai.Chiba. Prefectural Sanbu Regional Branch Office, 1-11, Higasishinshuku, Togane, Chiba 283-0006, Japan. h.asi2@pref.chiba.lg.jp
Keywords: Barremian; Choshi Group; Japan; Lower Cretaceous; marine fish; otolith
Final citation: Miyata, Shinya, Isaji, Shinji, Kashiwagi, Kenji, and Asai, Hidehiko. 2024. The first record of Lower Cretaceous otoliths from the Kimigahama Formation (Barremian) of the Choshi Group, Chiba Prefecture, Japan. Palaeontologia Electronica, 27(1):a20.
https://doi.org/10.26879/1318
palaeo-electronica.org/content/2024/5171-otoliths-from-cretaceous
Copyright: April 2024 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Fossil otoliths are important for understanding the paleobiogeography, paleoecology, and systematics of fishes. Fossil records of these otoliths are known from the Devonian through the Mesozoic to Holocene worldwide (e.g., Nolf, 1985). Numerous paleontological studies have been conducted on fossil otoliths from Cenozoic deposits (e.g., Schwarzhans, 2003, 2012; Girone and Nolf, 2009; Mitsui et al., 2021). Although Mesozoic otolith records are less abundant than Cenozoic records (e.g., Nolf, 1985; Schwarzhans, 2018), they offer a wealth of information, including biostratigraphic, paleobiogeographic, and paleoecological insights, as well as valuable details regarding the evolutionary history of fishes. The oldest otolith records are from the Lower Devonian-lower Middle Devonian (e.g., the Albanov Formation and Wood Bay Formation) (Nolf, 1985). Schwarzhans (2018) reviewed Jurassic-Lower Cretaceous otoliths to discuss the stratigraphic ranges of some actinopterygians with reference to the skeletal record.
Focusing on the fossil records of Cretaceous otoliths, some marine and nonmarine otoliths have been documented in Upper Cretaceous deposits. Nonmarine teleost otoliths have been reported in India (Rana, 1988, 1990, 1996; Rana and Shani, 1989; Nolf et al., 2008), while marine teleost otoliths have been found predominantly in North America and to a lesser extent in Europe and Asia (e.g., Yokoi, 1998; Stringer et al., 2016, 2020; Stringer and Schwarzhans, 2021; Schwarzhans and Stringer, 2020; Schwarzhans and Jagt, 2021, 2022; Stringer and Sloan, 2023) (Table 1). In contrast, assemblages of Lower Cretaceous marine fish otoliths have been primarily documented in the western Tethys area, with only a few reported in the USA. (Table 1). 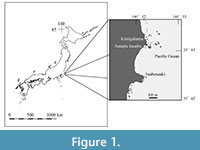 Limited findings on marine fossil actinopterygian fishes have been reported from Lower Cretaceous strata in East Asia. Specifically, only pachycormid? teeth have been described from the Lower Sebayashi Formation (Barremian) of the Sanchu Group. The depositional setting of the Lower Sebayashi Formation was likely a brackish water environment (Takakuwa et al., 2011). Thus, the fossil record of Early Cretaceous marine teleost fishes in East Asia is poorly represented. Recently, we discovered some marine fish otoliths in the shell bed of the Lower Cretaceous Kimigahama Formation, Choshi Group, Chiba Prefecture, Japan (Figure 1). This is the first record of marine fish otoliths from Lower Cretaceous strata in East Asia and provides valuable information about marine fish fauna and paleobiogeography in East Asia during the Early Cretaceous.
Limited findings on marine fossil actinopterygian fishes have been reported from Lower Cretaceous strata in East Asia. Specifically, only pachycormid? teeth have been described from the Lower Sebayashi Formation (Barremian) of the Sanchu Group. The depositional setting of the Lower Sebayashi Formation was likely a brackish water environment (Takakuwa et al., 2011). Thus, the fossil record of Early Cretaceous marine teleost fishes in East Asia is poorly represented. Recently, we discovered some marine fish otoliths in the shell bed of the Lower Cretaceous Kimigahama Formation, Choshi Group, Chiba Prefecture, Japan (Figure 1). This is the first record of marine fish otoliths from Lower Cretaceous strata in East Asia and provides valuable information about marine fish fauna and paleobiogeography in East Asia during the Early Cretaceous.
GEOLOGICAL SETTING
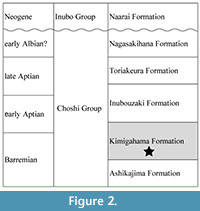 The Lower Cretaceous Choshi Group is distributed along the eastern coast of the Choshi Peninsula, Chiba Prefecture, Japan. The lithological division of the Choshi Group was first defined by Obata et al. (1975) but was subsequently reexamined by Obata et al. (1982). According to Obata et al. (1982), the Choshi Group is composed of the Ashikajima Formation (Barremian), Kimigahama Formation (Barremian-early Aptian), Inubouzaki Formation (early-late Aptian), Toriakeura Formation (late Aptian), and Nagasakihana Formation (early Albian), in ascending order (Obata et al., 1982; Obata and Matsukawa, 2007, 2009a, b) (Figure 2). Marine fish otoliths were collected from the lower horizon of the Kimigahama Formation. The Kimigahama Formation mainly consists of mudstone and alternating sandstone and mudstone (Obata et al., 1975, 1982; Obata and Matsukawa, 2007, 2009a, b). Katsura et al. (1984) and Ito and Matsukawa (1997) reported that the Kimigahama Formation consists of bioturbated silty sandstone, sandy siltstone, and hummocky and swaley cross-stratified sandstone deposited in an offshore to shoreface environment. This formation contains various kinds of faunal and floral taxa, including foraminifers, radiolarians, ammonoids, bivalves, gastropods, and palynomorphs (e.g., Obata et al., 1975, 1982; Hayami and Oji, 1980; Kase and Maeda, 1980; Obata and Matsukawa, 2007, 2009a, b; Legrand et al., 2011; Ando et al., 2014; Isaji et al., 2022). Obata et al. (1982) and Obata and Matsukawa (2009a) reported the presence of Hamulina, Heteroceras, Holcodiscus, Parasynoceras, Pulchellia, and Silesites ammonoids and Ammobaculites reophacoides, Epistomina hechti, Lenticulina heiermanni, Trochammina neocomiana, Verneuilinoides subfiliformis, and V. plexus neocomiensis foraminifera, and this fossil assemblage indicates that the Kimigahama Formation is Barremian in age (Obata and Matsukawa, 2009a).
The Lower Cretaceous Choshi Group is distributed along the eastern coast of the Choshi Peninsula, Chiba Prefecture, Japan. The lithological division of the Choshi Group was first defined by Obata et al. (1975) but was subsequently reexamined by Obata et al. (1982). According to Obata et al. (1982), the Choshi Group is composed of the Ashikajima Formation (Barremian), Kimigahama Formation (Barremian-early Aptian), Inubouzaki Formation (early-late Aptian), Toriakeura Formation (late Aptian), and Nagasakihana Formation (early Albian), in ascending order (Obata et al., 1982; Obata and Matsukawa, 2007, 2009a, b) (Figure 2). Marine fish otoliths were collected from the lower horizon of the Kimigahama Formation. The Kimigahama Formation mainly consists of mudstone and alternating sandstone and mudstone (Obata et al., 1975, 1982; Obata and Matsukawa, 2007, 2009a, b). Katsura et al. (1984) and Ito and Matsukawa (1997) reported that the Kimigahama Formation consists of bioturbated silty sandstone, sandy siltstone, and hummocky and swaley cross-stratified sandstone deposited in an offshore to shoreface environment. This formation contains various kinds of faunal and floral taxa, including foraminifers, radiolarians, ammonoids, bivalves, gastropods, and palynomorphs (e.g., Obata et al., 1975, 1982; Hayami and Oji, 1980; Kase and Maeda, 1980; Obata and Matsukawa, 2007, 2009a, b; Legrand et al., 2011; Ando et al., 2014; Isaji et al., 2022). Obata et al. (1982) and Obata and Matsukawa (2009a) reported the presence of Hamulina, Heteroceras, Holcodiscus, Parasynoceras, Pulchellia, and Silesites ammonoids and Ammobaculites reophacoides, Epistomina hechti, Lenticulina heiermanni, Trochammina neocomiana, Verneuilinoides subfiliformis, and V. plexus neocomiensis foraminifera, and this fossil assemblage indicates that the Kimigahama Formation is Barremian in age (Obata and Matsukawa, 2009a).
MATERIALS AND METHODS
The fossil otoliths described in the present study were obtained from a sandstone boulder (40×36×49 cm), which was sampled by Mr. Yamada, a local paleontologist, on May 5, 1998, and this boulder was adjacent to locality 7316 in Obata et al. (1982) (=Loc. 2 in Kase and Maeda, 1980; Figure 1). The boulder is derived from the Kimigahama Formation, which is distributed near locality 7316. It consists of a lenticular mollusk-rich sandstone bed, which is characteristic facies of the Kimigahama Formation (Isaji et al., 2022). These assemblages are identical to the matrices used in the studies of microgastropod assemblages by Isaji et al. (2022) and radiolarians (Ando et al., 2014).
The matrix was chemically dissolved by using the sodium tetraphenylborate method of Noda and Jin (2004). Various kinds of microfauna were recovered from the sandstone boulder: gastropods, bivalves, ammonoids, echinoderms, otoliths, foraminifers, and radiolarians. Among them, fossil otoliths were picked under a stereomicroscope by Isaji, S. Observations of otoliths were conducted under a stereomicroscope. The photographs were taken with a digital camera (Nikon D300), and the interpretative drawings were drawn directly using Photoshop 6.0 and an LCD pen tablet (Wacom Cintiq 12WX) during observation under a stereomicroscope. The otolith terminology followed that of Nolf (1985), and the fish classification followed that of Nelson et al. (2016). The specimens were assigned the prefix CBM-PV and stored at the Natural History Museum and Institute, Chiba.
RESULTS
An Otolith Assemblage from the Kimigahama Formation and Its Preservation
Otoliths from the Kimigahama Formation represent actinopterygian fauna. The following taxa were identified from seven specimens from Teleostei fam., gen. et sp. indet.; 83 specimens from Pterothrissinae; 17 specimens from Elopiformes gen. et sp. indet.; 32 specimens from Argentinidae gen. et sp. indet.; and seven specimens from Ichthyotringidae fam., gen. et sp. indet. (Table 2). Small otoliths, approximately 1 mm in size, such as Pterothrissinae gen. et sp. indet. are abundant. Additionally, while they retain their external form, many have a worn surface, and lack anterior parts. For example, the wavy external features of the shape of the otoliths of the family Argentinidae are obscured. In Ichthyotringidae, the striations and wavy external features of the shape are unclear. Therefore, we could not classify the species in more detail than at the subfamily level in this study.
SYSTEMATIC PALEONTOLOGY
Subclass ACTINOPTERYGII sensu Goodrich, 1930.
Subdivision TELEOSTEI Müller, 1845
Teleostei fam., gen. et sp. indet.
(Figure 3A-B)
 Reference material. CBM (Natural History Museum and Institute, Chiba)–PV 8203 (Figure 3A) and six other otoliths
Reference material. CBM (Natural History Museum and Institute, Chiba)–PV 8203 (Figure 3A) and six other otoliths
Description. The otoliths are thick, large, and approximately 3–5 mm length. The shape is nearly elliptical, the dorsal rim is straight, and the ventral and posterior rims are rounded. The interior side is convex. The dorsal margin of the posterior part of the rostral region is ventrally concave and slightly angulated. The margin is smooth and has no sculpturing. The sulcus is closed at the posterior end, and the ostium is open. The cauda is medial, straight, tubular, and elongated toward the posterior part. The anterior part of the rostrum is absent. The crista superior is present along the dorsal margin of the cauda, but the crista inferior is not recognizable.
Remarks. The present specimen is somewhat similar in the large and elliptical outline of sagitta with an elongated sulcus to that of Pterothrissinae such as Pteralbula galtina (see Schwarzhans, 2018, fig. 7E-G) or Pteralbula todolellana (Nolf, 2004, plate 2 fig. 1-6). However, the present specimen from the Barremian strata of the Kimigahama Formation exhibits a developed rostrum and straighter cauda. These characteristics distinguish it from typical Pterothrissinae. On the other hand, the specimen has a sulcus morphology that is like that of Argentinidae but differs from that of typical Argentinidae due to its larger size, reaching up to 5 mm, and lack of a pentagonal outer line.
The presence of Argentinidae and Pterothrissinae in the Valanginian strata of the Wawal claypit in Poland (Pindakiewicz et al., 2022) is consistent with the fact that these taxa are also present in the Barremian deposits of the Kimigahama Formation. Therefore, we suggest that the specimen represents either an Argentinidae or a Pterothrissinae. However, the specimen exhibits wear, making it challenging to definitively distinguish between the two groups. As a result, we provisionally assign it to Teleostei fam., gen. et sp. indet.
Superorder ELOPOMORPHA Müller, 1846
Order ALBULIFORMES Greenwood et al., 1966
Family ALBULIDAE Bleeker, 1859
Subfamily PTEROTHRISSINAE Greenwood, 1977
Pterothrissinae gen. et sp. indet.
(Figure 3C-E)
Reference material. CBM–PV 8205 (Figure 3C) and 82 other otoliths
Description. The otoliths are thick, small, and approximately 1 mm length. The shape is nearly elliptical, the dorsal rim is almost straight, and the ventral and posterior rims are rounded. The external surface is convex. The ostial region is funnel-shaped, and the rostrum is slightly peaked. The sulcus is closed at the posterior end, and the anterior opening is ostial. The cauda is medial, straight, and elongated toward the posterior part. The crista superior and crista inferior are developed along the dorsal and ventral margins of the cauda. The dorsal depression is present above the crista superior.
Remarks. The present specimens are characterized by their thickness, a slightly pointed trostrum, a rounded outline, and a cauda that does not extend to the posterior rim. It is like the small otolith of the Pteralbula galtina (Koken, 1891) of the Albian Folkstone Gault, England, which is an otolith-based species (Stinton., 1973; Schwarzhans, 2018). However, the present specimen is smaller (approximately 1 mm), and the external side is convex.
Otoliths of Pterothrissinae typically exceed 1 cm (e.g., Schwarzhans and Stringer, 2020; Schwarzhan and Jagt, 2021; Schwarzhans et al., 2022; Stringer and Sloan, 2023). Therefore, the otoliths of Pterothrissinae in the Kimigahama Formation may be juvenile fishes. The known Early Cretaceous Pterothrissinae species include Pteralbula galtina, P. cantina, P. todolellana, P. polonica, and Elopothrissus pawpawensis (Shepherd, 1916; Stinton, 1973; Nolf, 2004; Schwarzhans et al., 2022; Pindakiewicz et al., 2022; see Table 3). In the Early Cretaceous, Nolf (2004) described fossil otoliths of Pterothrissinae approximately 1 mm in length as the “genus Pterothrissidarum” sp. According to his study, it remains to be determined whether “genus Pterothrissidarum” sp. represents an individual variation of P. todolellana or a member of another species. Even in our otoliths of Pterothrissinae, it is challenging to determine whether these otoliths to a previously described species or a new species of Cretaceous Pterothrissinae. Therefore, in this study, otoliths of juvenile Pterothrissinae from the Kimigahama Formation were assigned to Pterothrissinae gen. et sp. indet.
Order ELOPIFORMES Sauvage, 1875
Elopiformes fam., gen. et sp. indet.
(Figure 4A-C)
 Reference material. CBM–PV 8206 (Figure 4A) and 16 other otoliths
Reference material. CBM–PV 8206 (Figure 4A) and 16 other otoliths
Description. The otoliths are thin, small, and approximately 2–3 mm length. The shape is elliptical, the dorsal rim is slightly convex, the posterior rim is rounded, and the ventral rim is straight. The external surface is flat. The ostium is funnel-shaped, and the rostrum is well-developed and pointed. The excisura is not notched. The sulcus is closed at the posterior end, and the anterior opening is ostial. The cauda is medial, straight, and elongated toward the posterior part and does not reach the posterior margin. The crista superior is developed along the dorsal margin of the cauda. There is no dorsal depression above the crista superior.
Remarks. The specimen is like the Protoelops scalpellum (Nolf, 2004) from the Aptian Forcall Formation, which is an otolith-based species. Protoelops scalpellum was assigned to be “Protacanthopterygian” in Nolf (2004); however, Schwarzhans (2018) assigned “Protacanthopterygiorum” to Protoelops, which belongs to Elopiformes. In our specimens, a thin, elliptical rostrum is developed, and the cauda does not reach the posterior rim. However, its dorsal rim is longer than that of the genus Protoelops. However, most specimens are fragmented, making it challenging to determine whether they can be attributed to a distinct genus or species. Therefore, in the present study, this species is assigned to Elopiformes gen. et sp. indet.
Indeterminate PROTACANTHOPTERYGII Greenwood et al., 1966
Order ARGENTINIFORMES Bertelsen 1958
Family ARGENTINIDAE Bonaparte 1846
Argentinidae gen. et sp. indet.
(Figure 4D-F)
Reference material. CBM–PV 8207 (Figure 4D) and 31 other otoliths
Description. The otoliths are thin and approximately 2–3 mm length. The shape is nearly pentagonal, the dorsal rim is almost straight, the ventral rim is sinuate, and the posterior rim is slightly angled. The ostium is funnel-shaped, and the rostrum is blunt. The sulcus is open at the posterior end, and the anterior opening is ostial. The cauda is medial, straight, and elongated toward the posterior end. The crista inferior is not well-delineated. The dorsal depression is present above the crista superior.
Remarks. The present specimen is like the Aptian species of Argentina? bergantinum (Nolf, 2004) because the shape is pentagonal in outline, the cauda is elongated toward the posterior end, and the ventral rim is sinuate. However, the rostrum is less developed than that of A.? bergantinum. Maastrichtian species of A. voigti Schwarzhans, 2010, Albian species of “A”. lobata (Stinton, 1973), Aptian species Argentina? texana Schwarzhans et al., 2022, and Valanginian species Palaeoargentina plicata Pindakiewicz et al., 2022 are known as Cretaceous otolith-based species. The Early Cretaceous genus Argentina is at least tentative or questionable (see Schwarzhans, 2018); therefore, we treated the present specimen as an undetermined species of the Argentinidae gen. et sp. indet. On the other hand, according to Schwarzhans (2018), these similar Argentinid otolith characteristics are plesiomorphic and have also been observed in extinct stem teleosts. The genus Leptolepis, which includes stem teleosts, also has a similar otolith morphology to that of Cretaceous otolith-based Argentinid species. However, the stratigraphic range of the Leptolepis or Leptolepis -type otoliths is Jurassic to Hauterivian, and most otolith-based species are from the Jurassic (Schwarzhans, 2018). Therefore, otoliths with shapes like those of the Cretaceous genus Argentina may need to be re-examined and/or discovered as otoliths in situ with the skeleton.
Order AULOPIFORMES Rosen, 1973
Suborder ICHTHYOTRINGOIDEI Goby, 1969
Family ICHTHYOTRINGIDAE Jordan, 1905
Ichthyotringidae gen. et sp. indet.
(Figure 4G)
Reference material. CBM–PV 8208 (Figure 4G) and 6 other otoliths
Description. The otoliths are slender, thin, and approximately 2 mm length. The dorsal and ventral rims are abraded, and the posterior rims are slightly rounded. Striations are observed on the inner surface of the ventral area. The ostial region is tubular, and the anterior part of the rostrum is missing. The sulcus is closed at the posterior end, and the anterior opening is ostial. The cauda is medial, straight, and elongated toward the posterior part, but does not reach the posterior margin. The crista superior and crista inferior are present along the margin of the cauda. The dorsal depression is present above the crista superior.
Remarks. The specimens reported here are similar to those of Ichthyotringidae, including Thrax acutus Stringer et al., 2020, Ichthyotringa? tavernei (Nolf and Stringer, 1996), I.? cuneata Schwarzhans et al., 2022, Apateodus crenellatus Schwarzhans and Stringer, 2020, Apateodus corneti (Forir, 1887), Apateodus ? assisi Schwarzhans et al., 2018b, and Apateodus sp. (Schwarzhans et al., 2018a, figures 7D-G) (Table 3). The present specimens have several characteristics in common with Ichthyotringa tavernei; the otolith is slender; the cauda is straight, long, and elongated toward the posterior part; and there are striations on the ventral area. However, the present specimens have a dorsal depression and are small (Ichthyotringa ? tavernei has no clear dorsal depression and reaches sizes greater than 3 mm). In I.? cuneata, the dorsal rim is clearly different from that of our specimen because it is square. Due to the wear and dissolution of this specimen, nothing more could be determined. Therefore, in the present study, these elongated otoliths were assigned to Ichthyotringidae, gen. et sp. indet.
DISCUSSION
Stratigraphical and Paleobiogeographical Distributions
This study describes an otolith assemblage consisting of four orders (Elopiformes, Albuliformes, Argentiniformes, and Aulopiformes). We focus on Pterothrissinae (Albuliformes), Elopiformes, Argentinidae (Argentiniformes), and Ichthyotringoidei (Aulopiformes). In previous studies of Cretaceous fossil records, these taxa are summarized in Table 3, and their occurrence locations are plotted on a paleomap in Figure 5 and Figure 6.
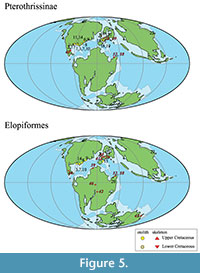 Pterothrissinae. The otoliths of the Cretaceous Pterothrissinae mainly existed from the Valanginian to late Maastrichtian in North America and Europe. The occurrences of the Lower Cretaceous specimens were limited to Europe and Texas, and the oldest specimens were from Valanginian strata in Poland (Stinton, 1973; Nolf, 2004; Schwarzhans, 2018b; Pindakiewicz et al., 2022; Schwarzhans et al., 2022). Skeleton-based specimens have been reported in Germany, Lebanon, and Mexico, and the oldest species is Nunaneichthys mexicanus, which is from the El Doctor Formation (Albian to Cenomanian) (Hernandez-Guerrero et al., 2020). The Pterothrissinae from the Kimigahama Formation is the first recorded Barremian specimen, and it fills a stratigraphic gap. According to Barros-García et al. (2018), the estimated divergence time between the genus Albula (Albulidae) and Pterothrissus (Pterothrissinae) is approximately 120 Ma (Aptian), according to a molecular phylogenetic study, which is supported by the Lower Cretaceous specimens described here.
Pterothrissinae. The otoliths of the Cretaceous Pterothrissinae mainly existed from the Valanginian to late Maastrichtian in North America and Europe. The occurrences of the Lower Cretaceous specimens were limited to Europe and Texas, and the oldest specimens were from Valanginian strata in Poland (Stinton, 1973; Nolf, 2004; Schwarzhans, 2018b; Pindakiewicz et al., 2022; Schwarzhans et al., 2022). Skeleton-based specimens have been reported in Germany, Lebanon, and Mexico, and the oldest species is Nunaneichthys mexicanus, which is from the El Doctor Formation (Albian to Cenomanian) (Hernandez-Guerrero et al., 2020). The Pterothrissinae from the Kimigahama Formation is the first recorded Barremian specimen, and it fills a stratigraphic gap. According to Barros-García et al. (2018), the estimated divergence time between the genus Albula (Albulidae) and Pterothrissus (Pterothrissinae) is approximately 120 Ma (Aptian), according to a molecular phylogenetic study, which is supported by the Lower Cretaceous specimens described here.
Elopiformes. The stratigraphic range of Elopiformes is from the Jurassic to recent (see Schwarzhans, 2018). Throughout the Cretaceous, both otoliths and skeletons of this group have been recorded in numerous fossil occurrences (see Table 3), especially the wide geographical distribution in the Northern and Southern Atlantic and Tethys Sea regions in the Early Cretaceous, but our records are limited to the Northern Atlantic and Western Tethys regions in the Late Cretaceous period (Figure 5, Table 3). Cavin (2008) examined the patterns of vicariance and dispersal in various groups of bony fishes. His study revealed that the east‒west pattern is more prevalent than the north‒south pattern in terms of the vicariance and dispersal patterns of Cretaceous bony fishes. Although assigning otoliths from the Kimigahama Formation to genus or species-level classifications is challenging, our study suggests an east-west dispersal pattern for this group during the Cretaceous. This pattern may indicate the radiation of the group not only toward the North Atlantic-Tethys region but also toward East Asia. However, the specific dispersal routes of Elopiformes in Cretaceous strata in East Asia have not been determined. Further data, including skeletal remains and otoliths, are needed to address this matter.
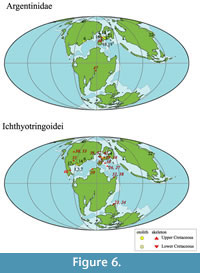 Argentinidae. Cretaceous Argentinidae have been described from the Valanginian to the late Maastrichtian in North America, Africa, and Europe. The oldest known species among the Argentinidae is Palaeoargentina plicata (otolith-based species), described from Valanginian deposits in Poland (Pindakiewicz et al., 2022). For the records on skeletal fossils, Nybelinoides pattersonella of the family Argentinidae was described from the Barremian to Aptian strata in Belgium (Taverne, 1982; 1999). These fossil records of Cretaceous argentinid fishes were limited to the North Atlantic region, mainly in Europe (Table 3). The argentinid otoliths from the Kimigahama Formation indicate that early argentinid fishes were also distributed in the Western Pacific Ocean during the Early Cretaceous (Figure 6).
Argentinidae. Cretaceous Argentinidae have been described from the Valanginian to the late Maastrichtian in North America, Africa, and Europe. The oldest known species among the Argentinidae is Palaeoargentina plicata (otolith-based species), described from Valanginian deposits in Poland (Pindakiewicz et al., 2022). For the records on skeletal fossils, Nybelinoides pattersonella of the family Argentinidae was described from the Barremian to Aptian strata in Belgium (Taverne, 1982; 1999). These fossil records of Cretaceous argentinid fishes were limited to the North Atlantic region, mainly in Europe (Table 3). The argentinid otoliths from the Kimigahama Formation indicate that early argentinid fishes were also distributed in the Western Pacific Ocean during the Early Cretaceous (Figure 6).
Ichthyotringoidei. Ichthyotringoidei includes two families, Ichthyotringidae and Apateopholidae, along with one indeterminate family, Ursichthys (Gody, 1965; Newbrey and Konishi, 2015; Silva and Gallo, 2011). Late Cretaceous Ichthyotringoidei otoliths have been documented in various formations, including the Ripley Formation, Severn Formation, Arkadelphia Formation and Owl Creek Formation in the USA, as well as the Maastricht Formation in the Netherlands. They are also present in the Lower Cretaceous Pawpaw Formation (Albian) (e.g., Stringer et al., 2020; Stringer and Schwarzhans, 2021; Schwarzhans and Jagt, 2021; see Table 3). In contrast, numerous skeletal fossils of Ichthyotringoidei, such as Apateodus, Apateopholis, Ursichthys and Ichthyotringa, have been recorded in the United States, Europe, and Morocco (e.g., Silva and Gallo, 2011; Newbrey and Konishi, 2015) (Table 3). These skeletal and otolith fossil records range from the Albian to the Maastrichtian. However, our study of the otolith fossils of Ichthyotringidae from the Kimigahama Formation indicates that the presence of Ichthyotringoidei dates to the Barremian. Additionally, fossil records of this family are concentrated mainly in the United States and the Tethys Sea region. Our findings suggest that the distribution of Ichthyotringoidei expanded there into East Asia during the Barremian. Newbrey and Konishi (2015) compiled a Cretaceous-Paleogene range chart for the Aulopiformes. According to their work, Aulopiformes fossils from the Barremian are also scarce, with only Acrognathus, Atolvorator, and an undetermined Aulopiformes species known, and no records during the Aptian. The absence of fossil records for Aptian Aulopiformes signifies a gap in the stratigraphic fossil record. Therefore, the dispersal routes that occurred during the Lower Cretaceous remain uncertain, emphasizing the importance of future studies.
Special Remarks on the Paleoecology of Cretaceous Pterothrissinae and Argentinidae
The recent species of Pterothrissinae and Argentinidae are deep sea-adapted teleosts. According to Hidaka et al. (2016), living Pterothrissinae consists of only two species, namely, Pterothrissus gissu, which is distributed at depths of 147–1000 m in the benthopelagic zone in China, Japan, and Russia (e.g., Aizawa, 2000; Sheiko and Fedorov, 2000; Aizawa and Doiuchi, 2013), and Nemoossis belloci, which is distributed at depths of 20–500 m (usually 100–400 m) in the bathydemersal zone from Mauritania to Sandwich Harbor, Namibia (Whitehead, 1981; Whitehead, 1990). Living Argentinidae are distributed in the benthopelagic zone on the outer shelf and upper slope, rarely to a depth of 1400 m (Paxton and Cohen, 1999).
Pterothrissinae and Argentinidae, which currently inhabit the deep sea, were obtained from Cretaceous shallow marine deposits (Table 3). The otoliths of Cretaceous Pterothrissinae mostly occur in shelf or shallower marine deposits, and deep marine or open ocean material is also known from only the Maastrichtian Severn Formation (Maryland, USA) and Gerhartsreiter Formation (Bavaria, Germany) (Huddleston and Savoie, 1983; Schwarzhans, 2010; Stringer and Schwarzhans, 2021). The Pterothrissinae from the Severn Formation and Gerhartsreiter Formation may be the oldest otoliths of this subfamily from the deep sea or open ocean deposits. On the other hand, a few skeleton-based fossils of Pterothrissinae are known from both the Lower and Upper Cretaceous strata and occur in deeper marine deposits, (Table 3). The otoliths of Pterothrissinae from the shore deposits of the Kimigahama Formation are consistent with those of previous studies. Thus, our study suggests that early Pterothrissinae may have expanded its relatively shallow marine habitats.
The otoliths of Cretaceous Argentinidae occur in the Lower Cretaceous shelf or shallower deposits, not only in shallow marine environments but also in Upper Cretaceous deep marine and open ocean deposits (see Table 3). The skeleton-based fossils of Argentinidae in Lower Cretaceous strata are known from lacustrine to outer sea shelf deposits, and Upper Cretaceous fossils occur in outer shelf marine deposits (see Table 3). The Valanginian deposits of the Wawal claypit, Poland, from which the earliest Argentinidae, Palaeoargentina plicata, was described, are interpreted as representing a shallow-water environment within the littoral zone (Pindakiewicz et al., 2022). Considering that the otolith of argentinid fish from the shore deposit of the Kimigahama Formation is also one of the early records in this family, the early Argentinidae inhabited shallow marine and nonmarine environments. Therefore, it is probable that the habitats of Argentinidae were nonmarine to shallow-water environments in the Early Cretaceous and shifted to a deeper environment during the Late Cretaceous.
CONCLUSION
We recognized a total of five otolith types: undetermined Teleostei, Pterothrissinae, Elopiformes, Argentinidae, and Ichthyotringidae. Our new data on these otoliths from Barremian strata in the northwestern Pacific indicate that these taxa were widely distributed in the Pacific, Tethys, and Atlantic seas during the Early Cretaceous. Specifically, because of the adaptation of living Argentinidae and Pterothrissinae to deep marine environments, these taxa are also found in Lower Cretaceous deposits, such as the Kimigahama Formation, which indicates a shallow marine depositional setting. Fossils of these groups in deep-sea sediments have been found in at least Late Cretaceous strata in previous studies. Therefore, the occurrence of otolith fossils of these “deep-sea fishes” in Early Cretaceous shallow marine sediments seems to be consistent with the findings of previous studies. In addition, otoliths from the Kimigahama Formation contributed to our understanding of the ecology of “deep-sea fish” before they adapted to the deep sea.
ACKNOWLEDGMENTS
The authors wish to express their sincere gratitude to Y. Katsuhiko (Choshi City) for collecting the fossil-bearing sandstone and donating it to the Natural History Museum and Institute, Chiba.
The authors are deeply grateful to A. Hisao (Ibaraki University) for providing valuable advice to the Choshi Group.
The authors are also deeply grateful to H. Tischlinger (Jura-Museum Eichstätt), T.L. Harrell (Geological Survey of Alabama), and L. Le Callonnec (Sorbonne University). for providing references. The authors express their sincere gratitude to W.W. Schwarzhans (Zoological Museum, Natural History Museum of Denmark) and C.H. Lin (Biodiversity Research Center, Academia Sinica), who reviewed this manuscript and provided valuable comments and suggestions that helped to improve the final version.
This study was partly supported by Grants-in-Aid for Scientific Research (KAKENHI) from the Japan Society for the Promotion of Science to S. Isaji (no. 19654077, 2007–2009) and K. Kashiwagi (no. 23540547, 2011–2013).
REFERENCES
Aizawa, M. 2000. Pterothrissidae, p. 190, 1451. In Nakabo, T. (ed.), Fishes of Japan with pictorial keys to the species, second edition. Tokai University Press, Tokyo. [In Japanese]
Aizawa, M. and Doiuchi, R. 2013. Pterothrissidae, p. 236, 1781. In Nakabo, T. (ed.), Fishes of Japan with pictorial keys to the species, third edition. Tokai University Press, Hadano. [In Japanese)]
Albrich, S., Frijia, G., Parente, M., and Caus, E. 2014. The evolution of the earliest representatives of the genus Orbitoides : implications for Upper Cretaceous biostratigraphy. Cretaceous Research, 51:22–34.
https://doi.org/10.1016/j.cretres.2014.04.013
Albrich, S., Boix, C., and Caus, E. 2015. Selected agglutinated larger foraminifera from the Font de les Bagasses unit (Lower Campanian, Southern Pyrenees). Carnets de Geologie, 15:245–267.
https://doi.org/10.4267/2042/57953
Allen, J.G. and Shimada, K. 2021. Fossil vertebrates from a unique marine bonebed of the Upper Cretaceous Smoky Hill Chalk, western Kansas, USA: New insights into the paleoecology of the Niobrara Formation. Journal of Vertebrate Paleontology, 41:e2066999.
https://doi.org/10.1080/02724634.2021.2066999
Alves, Y.M., Alvarado-Ortega, J., and Brito, P.M. 2020. † Epaelops martinezi gen. and sp. nov. from the Albian limestone deposits of the Tlayúa quarry, Mexico-A new late Mesozoic record of Elopiformes of the western Tethys. Cretaceous Research, 110:104260.
https://doi.org/10.1016/j.cretres.2019.104260
Amalfitano, J., Giusberti, L., Fornaciari, E., and Carnevale, G. 2020. Upper Cenomanian fishes from the Bonarelli level (OAE2) of Northeastern Italy. Rivista Italiana di Paleontologia e Stratigrafia, 126:261–314.
https://doi.org/10.13130/2039-4942/13224
Ando, H., Kashiwagi, K., Hirayama, R., and Toshimitsu, S. 2014. Cretaceous forearc basin siliciclastic successions along the Pacific coast, central Japan: Choshi, Nakaminato and Futaba groups. Field Excursion Guide of the Second International Symposium of International Geoscience Programme (IGCP) Project 608, p. 1–42.
Argyriou, T. and Davesne, D. 2021. Offshore marine actinopterygian assemblages from the Maastrichtian-Paleogene of the Pindos Unit in Eurytania, Greece. PeerJ, 9:e10676.
https://doi.org/10.7717/peerj.10676
Argyriou, T., Alexopoulos, A.J. Carrillo-Briceño, D., and Cavin, L. 2022. A fossil assemblage from the mid-late Maastrichtian of Gavdos Island, Greece, provides insights into the pre-extinction pelagic ichthyofaunas of the Tethys. PLoS ONE, 17:e0265780.
https://doi.org/10.1371/journal.pone.0265780
Audo, D. and Charbonnier, S. 2012. Late Cretaceous crest-bearing shrimps from the Sahel Alma Lagerstätte of Lebanon. Acta Palaeontologica Polonica, 58:335–349.
https://doi.org/10.4202/app.2011.0056
Barros-García, D., Froufe, E., Bañón, R., Arronte, J.C., and de Carlos, A. 2018. Phylogenetic analysis shows the general diversification pattern of deep-sea notacanthiforms (Teleostei: Elopomorpha). Molecular Phylogenetics and Evolution, 124:192–198.
https://doi.org/10.1016/j.ympev.2018.03.007
Bartholomai, A. 2010. Revision of Flindersichthys denmeadi Longman 1932, a marine teleost from the Lower Cretaceous of the Great Artesian Basin. Queensland. Memoirs of the Queensland Museum, 55:43–68.
Butt, A. and Herm, D. 1978. Paleo-Oceanographic aspects of the upper cretaceous geosynclinal sediments of the Eastern Alps, p. 87–94. In Closs, H., Roeder, D., and Schmidt, K. (eds.), Alps, Apennines, Hellenides: Geodynamic Investigations Along Geotraverses by an International Group of Scientists, Schweizerbart’sche Verlagsbuchhandlung (Nägele u. Obermiller), Stuttgart.
Cavin, L. 2008. Palaeobiogeography of Cretaceous bony fishes (Actinistia, Dipnoi and Actinopterygii). Geological Society, London, Special Publications, 295:165–183.
https://doi.org/10.1144/SP295.11
Cochran, J.K., Landman, N.H., Turekian, K.K., Michard, A., and Schrag, D.P. 2003. Paleoceanography of the late Cretaceous (Maastrichtian) Western Interior seaway of North America: evidence from Sr and O isotopes. Palaeogeography, Palaeoclimatology, Palaeoecology, 191:45–64.
https://doi.org/10.1016/S0031-0182(02)00642-9
Corentin, P., Deconinck, J.F., Pellenard, P., Amédro, F., Bruneau, L., Chenot, E., Matrion, B., Huret, E., and Landrein, P. 2020. Environmental and climatic controls of the clay mineralogy of Albian deposits in the Paris and Vocontian basins (France). Cretaceous Research, 108:104342.
https://doi.org/10.1016/j.cretres.2019.104342
Cripps, J.A., Widdowson, M., Spicer, R.A., and Jolley, D.W. 2005. Coastal ecosystem responses to late stage Deccan Trap volcanism: the post K-T boundary (Danian) palynofacies of Mumbai (Bombay), west India. Palaeogeography, Palaeoclimatology, Palaeoecology, 216:303–332.
https://doi.org/10.1016/j.palaeo.2004.11.007
Dietze, K. 2009. Morphology and phylogenetic relationships of certain neoteleostean fishes from the Upper Cretaceous of Sendenhorst, Germany. Cretaceous Research, 30:559–574.
https://doi.org/10.1016/j.cretres.2008.11.001
Embry, J.C., Vennin, E., Van Buchem, F.S.P., Schroeder, R., Pierre, C., and Aurell, M. 2010. Sequence stratigraphy and carbon isotope stratigraphy of an Aptian mixed carbonate-siliciclastic platform to basin transition (Galve sub-basin, NE Spain). Geological Society, London, Special Publications, 329:113–143.
https://doi.org/10.1144/SP329.6
Fielitz, C. and González-Rodríguez, K.A. 2008. A new species of Ichthyotringa from the El Doctor Formation (Cretaceous), Hidalgo, Mexico, p. 373–388. In Arratia, G., Schultze, H.-P. and Wilson, M.V.H. (eds.), Mesozoic Fishes 4–Homology and Phylogeny, Verlag Dr. Friedrich Pfeil, München, Germany.
Fielitz, C. and Shimada, K. 2009. A new species of Apateodus (Teleostei: Aulopiformes) from the Upper Cretaceous Niobrara Chalk of western Kansas, U.S.A. Journal of Vertebrate Paleontology, 29:650–658.
https://doi.org/10.1671/039.029.0308
Fielitz, C. and Shimada, K. 2020. A possible undescribed aulopiform fish allied to the genus Apateodus from the Upper Cretaceous Niobrara Chalk of Kansas, U.S.A. Transactions of the Kansas Academy of Science, 123:435–440.
https://doi.org/10.1660/062.123.0313
Forey, P.L. 1973. A revision of the elopiform fishes, fossil and recent. Bulletin of the British Museum (Natural History), Geology, 10 (suppl.):1–222.
Forey, P.L., Lu, Y., Patterson, C., and Davies, C.E. 2003. Fossil fishes from the Cenomanian (Upper Cretaceous) of Namoura, Lebanon. Journal of Systematic Palaeontology, 1:227–330.
https://doi.org/10.1017/S147720190300107X
Frazier W.J. 1997. Upper Cretaceous strata in southwestern Georgia and adjacent Alabama. Atlanta Geological Society Field Trip Guidebook. Atlanta Geological Society, Atlanta.
Girone, A. and Nolf, D. 2009. Fish otoliths from the Priabonian (Late Eocene) of North Italy and South-East France-Their paleobiogeographical significance. Revue de Micropaléontologie, 52:195–218.
https://doi.org/10.1016/j.revmic.2007.10.006
Goody, P.C. 1969. The relationships of certain Upper Cretaceous teleosts with special reference to the myctophoids. Bulletin of the British Museum (Natural History), Geology Supplement, 7:1–259.
https://doi.org/10.1086/406502
Hattin, D.E. 1982. Stratigraphy and depositional environment of Smoky Hill Chalk Member, Niobrara Chalk (Upper Cretaceous) of the type area, western Kansas. Kansas Geological Survey, Bulletin 225:1–108.
Hayami, I. and Oji, T. 1980. Early Cretaceous bivalvia from the Chochi district, Chiba Prefecture, Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Series, 120:419–448.
Hernández-Guerrero, C., Cantalice, K.M., González-Rodríguez, K.A., and Bravo-Cuevas, V.M. 2020. The first record of a pterothrissin (Albuliformes, Albulidae) from the Muhi Quarry, mid-Cretaceous (Albian-Cenomanian) of Hidalgo, central Mexico. Journal of South American Earth Sciences, 103032.
https://doi.org/10.1016/j.jsames.2020.103032
Hidaka, K., Tsukamoto, Y., and Iwatsuki, Y. 2016. Nemoossis , a new genus for the eastern Atlantic long-fin bonefish Pterothrissus belloci Cadenat 1937 and a redescription of P. gissu Hilgendorf 1877 from the northwestern Pacific. Ichthyological Research, 64:45–53.
https://doi.org/10.1007/s10228-016-0536-5
Hilbrecht, H. and Dahmer, D.D. 1994. Sediment dynamics during the Cenomanian-Turonian (Cretaceous) oceanic anoxic event in Northwestern Germany. Facies, 30:63–83.
https://doi.org/10.1007/BF02536890
Hoganson, J., Erickson, J., and Holland, F. 2019. Chondrichthyan and osteichthyan paleofaunas from the Cretaceous (late Maastrichtian) Fox Hills Formation of North Dakota, USA: Paleoecology, Paleogeography, and Extinction. Bulletins of American Paleontology, 398:1–94.
Huddleston, R.W. and Savoie, K.M. 1983. Teleostean otoliths from the late Cretaceous (Maestrichtian age) Severn Formation of Maryland. Proceedings of the Biological Society of Washington, 96:658–663.
Isaji, S., Haga, T., and Kashiwagi, K. 2022. Early Cretaceous Small-Sized Gastropods from the Shallow Marine Deposits of the Kimigahama Formation, Choshi Group, Japan. Paleontological Research, 26:31–54.
http://doi.org/10.2517/PR20009
Ito, M. and Matsukawa, M. 1997. Diachronous evolution of third-order depositional sequences in the Early Cretaceous forearc basins: shallow marine and paralic successions in the Sanchu and Choshi Basins, Japan. Memoirs of the Geological Society of Japan, 48:60–75.
Jeans, C.V. 1980. Early submarine lithification in the Red Chalk and Lower Chalk of eastern England: a bacterial control model and its implications. Proceedings of the Yorkshire Geological Society, 43:81–157.
https://doi.org/10.1144/pygs.43.2.81
Kase, T. and Maeda, H. 1980: Early Cretaceous Gastropoda from the Choshi district, Chiba Prefecture, central Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Series, 118: 291–324.
Katsura, Y., Masuda, F., and Obata, I. 1984. Storm dominated shelf sea from the Lower Cretaceous Choshi Group, Japan. Annual Report of the Institute of Geoscience, the University of Tsukuba, 10:92–95.
Khalloufi, B., Ouarhache, D., and Leliévre, H. 2010. New paleontological and geological data about Jbel Tselfat (Late Cretaceous of Morocco). Historical Biology, 22:57–70.
https://doi.org/10.1080/08912961003668756
Knight, R.I. 1997. Benthic palaeoecology of the Gault Clay Formation (mid-and basal upper Albian) of the western Anglo-Paris Basin. Proceedings of the Geologists’ Association, 108:81–103.
https://doi.org/10.1016/S0016-7878(97)80032-6
Kocurek, G. 1978 Deltaic deposits in Kemp (Upper Cretaceous) and Kincaid (Paleocene) Formations, south-central Texas. Geological Society of America, South-Central Section, abstracts with program, 10:21.
Koken, E. 1891. Neue Untersuchungen an tertiären Fisch-otolithen II. Zeitschrift der deutschen geologischen Gesellschaft, 43:77–170.
Kriwet, J. and Gloy, U. 1995. Zwei mesopelagische Raubfische (Actinopterygii: Euteleostei) aus dem Unterturon der Kronsberg-Mulde bei Hannover/Misburg (NWDeutschland). Berliner Geowissenschaftliche Abhandlungen, Reihe E, 16:335–355.
Kruizinga, P. 1924. Apateodus corneti (For) in the Senonian beds of the southern part of Limburg (Netherland). Proceedings of the Science Section of the Koninklijke Nederlandse Akademie van Wetenschappen, 27:293–312.
Legrand, J., Pons, D., Nishida, H., and Yamada, T. 2011. Barremian palynofloras from the Ashikajima and Kimigahama formations (Choshi Group, Outer Zone of south-west Japan). Geodiversitas, 33:87–135.
https://doi.org/10.5252/g2011n1a6
López-Arbarello, A. 2004. The record of Mesozoic fishes from Gondwana (excluding India and Madagascar), p. 597–624. In Arratia, G. and Tintori, A. (eds.), Mesozoic Fishes 3–Systematics, Paleoenvironments and Biodiversity. Verlag Dr. Friedrich Pfeil, Munich.
Maisey, J.G. (ed.). 1991. Santana Fossils an Illustrated Atlas. T.F.H. Publications, Neptune City, New Jersey.
Mitsui, S., Taru, H., Ohe, F., Lin, C.H., and Strüssmann, C.A. 2021. Fossil fish otoliths from the Chibanian Miyata Formation, Kanagawa Prefecture, Japan, with comments on the paleoenvironment. Geobios, 64:47–63.
https://doi.org/10.1016/j.geobios.2020.11.003
Murray, A.M. 2000. The Palaeozoic, Mesozoic and Early Cenozoic fishes of Africa. Fish and Fisheries, 1:111–145.
https://doi.org/10.1046/j.1467-2979.2000.00015.x
Nelson, J.S., Grande, T.C., and Wilson, M.V.H. 2016. Fishes of the World (5th Edition). John Wiley & Sons, Inc., Hoboken, New Jersey.
https://doi.org/10.1002/9781119174844
Newbrey, M.G. and Konishi, T. 2015. A new lizardfish (Teleostei, Aulopiformes) from the Late Cretaceous Bearpaw Formation of Alberta, Canada, with a revised diagnosis of † Apateodus (Aulopiformes, Ichthyotringoidei). Journal of Vertebrate Paleontology 35:e918042.
https://doi.org/10.1080/02724634.2014.918042
Noda, Y. and Jin, X. 2004. Chemical treatment of dinosaur eggshells by using the sodium tetraphenylborate method. Memoir Fukui Prefectural Dinosaur Museum, 3: 39–46. [In Japanese with English abstract]
Nolf, D. 1985. Otolithi piscium. In Schultze, H. (ed.), Handbook of Paleoichthyology, 10. Gustav Fischer Verlag, Stuttgart.
Nolf, D. 2003. Fish otoliths from the Santonian of the Pyrenean faunal province, and an overview of all otolith-documented North Atlantic Late Cretaceous teleosts. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 73:155–173.
Nolf, D. 2004. Otolithes des poissons aptiens du Maestrazgo (province de Castellon, Espagne Orientale). Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, 74:101–120.
Nolf, D. 2016. Otoliths of Cenomanian fishes in the Ballon Marl (France), the earliest fossil record of perciform fishes. Bulletin d’ information des Géologues du Bassin de Paris, 53:14–20.
Nolf, D. and Dockery, D.T. 1990. Fish otoliths from the Coffee Sand (Campanian) of northeastern Mississippi. Mississippi Geology, 10:1–14.
Nolf, D. and Stringer, G.L. 1996. Cretaceous fish otoliths-a synthesis of the North American record, p. 433–459. In Arratia, G. and Viohl, G. (eds.), Mesozoic Fishes–Systematics and Paleoecology. Verlag Dr. Friedrich Pfeil, München, Germany.
Nolf, D., Rana, R.S., and Prasad, G.V. 2008. Late Cretaceous (Maastrichtian) fish otoliths from the Deccan intertrappean beds, India: a revision. Bulletin de l’Institut Royal des Sciences Naturelles de Belqique, Sciences de la Terre, 78:239–259.
Obata, I. and Matsukawa, M. 2007. Barremian-Aptian (Early Cretaceous) ammonoids from the Choshi Group, Honshu (Japan). Cretaceous Research, 28:363–391.
https://doi.org/10.1016/j.cretres.2006.06.004
Obata, I. and Matsukawa, M. 2009a. Supplementary description of the ammonoids from the Barremian to the Albian of the Choshi Peninsula, Japan. Cretaceous Research, 30:253–269.
https://doi.org/10.1016/j.cretres.2008.07.004
Obata, I. and Matsukawa, M. 2009b. Some ammonoids from the Barremian and probable Albian of the Choshi Peninsula, Japan. Bulletin of Tokyo Gakugei University. Natural Sciences, 61:97–103.
Obata, I., Hagiwara, S., and Kamiko, S. 1975. Geological age of the Cretaceous Choshi Group. Bulletin of the National Science Museum, C (Geology & Paleontology), 1:17–33.
Obata, I., Maiya, S., Inoue, Y., and Matsukawa, M. 1982. Integrated mega-and micro fossil biostratigraphy of the Lower Cretaceous Choshi Group, Japan. Bulletin of the National Science Museum, C (Geology & Paleontology), 8:145–179.
Oboh-Ikuenobe, F.E., Spencer, M.K., Campbell, C.E., and Haselwander, R.D. 2012. A portrait of Late Maastrichtian and Paleocene palynoflora and paleoenvironment in the northern Mississippi Embayment, southeastern Missouri. Palynology, 36:63–79.
https://doi.org/10.1080/01916122.2012.679208
Paxton, J.R. and Cohen, D.M. 1999. Argentinidae, Bathylagidae, Microstomatidae, Opisthoproctidae, p. 1884–1886. In Carpenter, K.E. and Niem, V.H. (eds.), The Living Marine Resources of the Western Central Pacific. FAO, Rome.
Pindakiewicz, M.K., Hryniewicz, K., and Kaim, A. 2022. Early Cretaceous radiation of teleosts recorded by the otolith-based ichthyofauna from the Valanginian of Wąwał, central Poland. Journal of Vertebrate Paleontology, 42:e2232008.
https://doi.org/10.1080/02724634.2023.2232008
Pollock, R.E. 1976. The depositional environments of the Maastricht and Kunrade Chalks (Maastrichtian) from the type area of Limburg, Netherlands. Staringia, 3:16–18.
Rana, R.S. 1988. Freshwater fish otoliths from the Deccan trap associated sedimentary (Cretaceous-Tertiary transition) beds of Rangapur, Hyderabad, District, Andhra Pradesh, India. Geobios, 21:465–493.
https://doi.org/10.1016/S0016-6995(88)80045-7
Rana, R.S. 1990. Palaeontology and Palaeoecology of the Intertrappean (Cretaceous-Tertiary transition) beds of the Peninsular India. Journal of the Palaeontological Society of India, 35:105–120.
Rana, R.S. 1996. Additional fish otoliths from the Deccan Trap associated sedimentary beds exposed near Rangapur, Rangareddi district, p. 477–492. In Pandey, J., Azmi, R.J., Bhandari, A., and Dave, A., (eds.), Contributions of the XV Indian Colloquium on Micropalaeontology and Stratigraphy, Wadia Institute of Himalayan Geology Dehradun, India.
Rana, R.S. and Sahni, A. 1989. Fish otoliths from the Takii Fonnation (Intertrappean Beds) of Nagpur, India. Geoscience Journal, 40:145–164.
Riegraf, W. 1995. Radiolarien, Diatomeen, Cephalopoden und Stratigraphie im pelagischen Campanium Westfalens (Oberkreide, NW-Deutschland). Neues Jahrbuch fur Geoiogie und Palaontologie, Abhandlungen 197:129–200.
https://doi.org/10.1127/njgpa/197/1995/129
Robaszynski, F., Gale, A., Juignet, P., Amédro, F., and Hardenbol, J. 1998. Sequence stratigraphy in the Upper Cretaceous series of the Anglo-Paris Basin: exemplified by the Cenomanian stage, p. 363–386. In de Graciansky, P.C., Hardenbol, J., Jacquin, T., and Vail, P.R. (eds.), Mesozoic and Cenozoic Sequence Stratigraphy of European Basins, SEPM Special Publication No. 60. SEPM Society for Sedimentary Geology, U.S.A.
Robert, P. and Yapaudjian, L. 1990. Early Cretaceous rift sediments of the Gabon-Congo Margin: lithology and organic matters; tectonic and paleogeothermal evolution. Journal of African Earth Sciences, 10:319–330.
https://doi.org/10.1016/0899-5362(90)90063-K
Schwarzhans, W. 1996. Otoliths from the Maastrichtian of Bavaria and their evolutionary significance, p. 417–431. In Arratia, G. and Viohl, G. (eds.). Mesozoic Fishes-Systematics and Paleoecology, Verlag Dr. Friedrich Pfeil, Munich, Germany.
Schwarzhans, W. 2003. Fish otoliths from the Paleocene of Denmark. Geological Survey of Denmark and Greenland Bulletin, 2:1–94.
Schwarzhans, W. 2010. Otolithen aus den Gerhartsreiter Schichten (Oberkeide: Maastricht) des Gerhartsreiter Grabens (Oberbayern). Palaeo Ichthyologica, 4:1–100.
Schwarzhans, W. 2012. Fish otoliths from the Paleocene of Bavaria (Kressenberg) and Austria (Kroisbach and Oiching-Graben). Palaeo Ichthyologica, 12:1–88.
Schwarzhans, W. 2018. A review of Jurassic and Early Cretaceous otoliths and the development of early morphological diversity in otoliths. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 287:75–121.
https://doi.org/10.1127/njgpa/2018/0707
Schwarzhans, W. and Jagt, J. 2021. Silicified otoliths from the Maastrichtian type area (Netherlands, Belgium) document early gadiform and perciform fishes during the Late Cretaceous, prior to the K/Pg boundary extinction event. Cretaceous Research, 127:104921.
https://doi.org/10.1016/j.cretres.2021.104921
Schwarzhans, W. and Jagt, J. 2022. Silicified bony fish otoliths from the Vaals Formation (lower Campanian) of Vaals-Eschberg, the Netherlands. Cretaceous Research, 139:105312.
https://doi.org/10.1016/j.cretres.2022.105312
Schwarzhans, W., and Milan, J. 2017. After the disaster: Bony fish remains (mostly otoliths) from the K/Pg boundary section at Stevns Klint, Denmark, reveal consistency with teleost faunas from later Danian and Selandian strata. Bulletin of the Geological Society of Denmark, 65:59–74.
https://doi.org/10.37570/bgsd-2017-65-05
Schwarzhans, W. and Stringer, G.L. 2020. Fish Otoliths from the Late Maastrichtian Kemp Clay (Texas, Usa) and the Early Danian Clayton Formation (Arkansas, Usa) and an Assessment of Extinction and Survival of Teleost Lineages Across the K-Pg Boundary Based on Otoliths. Rivista Italiana di Paleontologia e Stratigrafia, 126: 395–446.
https://doi.org/10.13130/2039-4942/13425
Schwarzhans, W., Beckett, H., Schein, J., and Friedman, M. 2018a. Computed tomography as a tool for linking the skeleton- and the otolith-based fossil records of teleost fishes. Palaeontology, 61:511–541.
https://doi.org/10.1111/pala.12349
Schwarzhans, W., Huddleston, R.W., and Takeuchi, G.T. 2018b. A Late Santonian fish-fauna from the Eutaw Formation of Alabama reconstructed from otoliths. Rivista Italiana di Paleontologia e Stratigrafia, 124:45–72.
https://doi.org/10.13130/2039-4942/9624
Schwarzhans, W., Stringer, G.L., and Welton, B. 2022. Oldest Teleostean Otolith Assemblage from North America (Pawpaw Formation, Lower Cretaceous, upper Albian, northeast Texas, USA). Cretaceous Research, 140:105307.
https://doi.org/10.1016/j.cretres.2022.105307
Scotese, C.R. 2014. Atlas of Late Cretaceous paleogeographic maps, PALEOMAP atlas for Arc GIS, volume 2, The Cretaceous, Maps 16–22, Mollweide Projection.
https://doi.org/10.13140/2.1.4099.4560
Sheiko B.A. and Fedorov, V.V. 2000. Part 1, p. 7–69. In Moiseev, R.S. and Tokranov, A.M. (eds.), Catalogue of the vertebrates of Kamchatka and adjacent waters. Kamchatskiy Petchatniy Dvor, Petropavlovsk-Kamchatsky. [In Russian]
Shepherd, C.E. 1916. Fossil otoliths. Knowledge, 39:177–184.
Sieber, R. and Weinfurter, E. 1967. Otolithen aus tiefen Gosauschichten Österreichs. Annalen des Naturhistorisches Museum Wien, 71:353–361.
Silva, H.M.A and Gallo, V. 2011. Taxonomic review and phylogenetic analysis of Enchodontoidei (Teleostei: Aulopiformes). Anais da Academia Brasileira de Ciências, 83:483–511.
https://doi.org/10.1590/S0001-37652011000200010
Spagna, P., Yans, J., Schnyder, J., and Dupuis, C. 2012. The paleoenvironment of the Bernissart Iguanodons: Sedimentological analysis of the Lower Cretaceous Wealden facies in the Bernissart area, p. 87–96. In Godefroit, P. (ed.), Bernissart dinosaurs and early terrestrial ecosystems, Indiana University Press, Indiana, U.S.A.
Stinton, F.C. 1973. Fish otoliths from the English Cretaceous. Palaeontology, 16:293–305.
Stringer, G.L. 1991. Upper Cretaceous (Maastrichtian) teleostean otoliths from the Ripley Formation, Union County, Mississippi. Mississippi Geology, 11:9–20.
Stringer, G.L. 2016. Late Cretaceous actinopterygians represented by otoliths from the Coon Creek Site in southwest Tennessee. Bulletin of the Alabama Museum of Natural History, 33:77–93.
Stringer, G.L. and Schwarzhans, W. 2021. Upper Cretaceous teleostean otoliths from the Severn Formation (Maastrichtian) of Maryland, USA, with an unusual occurrence of Siluriformes and Beryciformes and the oldest Atlantic coast Gadiformes. Cretaceous Research, 125:104867.
https://doi.org/10.1016/j.cretres.2021.104867
Stringer, G.L. and Sloan, J.C. 2023. First Cretaceous teleostean otolith assemblage (Arkadelphia Formation, upper Maastrichtian) from Arkansas, USA, early Gadiformes, and the Western Interior Seaway. PaleoBios, 40:1–39.
https://doi.org/10.5070/P940361192
Stringer, G.L., Oman, L.D., and Badger, R.F. 2016. Woodbury Formation (Campanian) in New Jersey yields largest known Cretaceous otolith assemblage of teleostean fishes in North America. Proceedings of the Academy of Natural Sciences of Philadelphia, 165:15–36.
https://doi.org/10.1635/053.165.0101
Stringer, G.L., Clements, D., Sadorf, E., and Shannon, K. 2018. First description and significance of Cretaceous teleostean otoliths (Tar Heel Formation, Campanian) from North Carolina. Eastern Paleontologist, 1:1–22.
Stringer, G.L., Schwarzhans, W., Phillips, G., and Lambert, R. 2020. Highly diversified late cretaceous fish assemblage revealed by otoliths (Ripley Formation and Owl Creek Formation, northeast Mississippi, USA). Rivista Italiana di Paleontologia e Stratigrafia, 126:111–155.
https://doi.org/10.13130/2039-4942/13013
Sutherland, J.T.F., Austen, P.A., Duffin, C.J., and Benton, M.J. 2017. Leptolepid otoliths from the Hauterivian (Lower Cretaceous) Lower Weald Clay (southern England). Proceedings of the Geologists’ Association, 128:613–625.
https://doi.org/10.1016/j.pgeola.2017.05.001
Takakuwa, Y., Sato, K., Kimura, T., and Kubota, K. 2011. Paleontological study of Sanchu Group. Natural history survey report of Gunma Museum of Natural History, 5:99–116. [In Japanese]
Taverne, L. 1975. Étude complémentaire de Wenzia congolensis (Arambourg C. & Schneegans, D., 1935), téléostéen salmoniforme fossile du Wealdien du Gabon, précédement décrit dans le genre Leptolepis Agassiz, L., 1832. Bulletin du Muséum National d’Histoire Naturelle, Science de la Terre, Troisième Série, 344:229–240.
Taverne, L. 1976a. Les téléostéens fossiles du Crétacé moyen de Kipala (Kwango, Zaïre). Annales du Musée Royal de l’Afrique Centrale, série in-8, Sciences Géologiques, Tervuren, 79:1–50.
Taverne, L. 1976b. Mutanda Ichthyologica Wenzichthys nomen novum en remplacement de Wenzia Taverne, L., 1975, téléostéen salmoniforme fossile du Crétacé inférieur du Gabon et de Guineé Équatoriale. Revue Zoologique Africaine, 90:1032.
Taverne, L. 1982. Sur Pattersonella formosa (Traquair, R.H. 1911) et Nybelinoides brevis (Traquair, R.H. 1911), Téléostéens Salmoniformes Argentinoïdes du Wealdien inférieur de Bernissart, Belgique, précédemment attribués au genre Leptolepis Agassiz, L. 1832. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, 54:1–27.
Taverne, L. 1999. Ostéologie et position systématique d’ Arratiaelops vectensis gen. nov., téléostéen élopiforme du Wealdien (Crétacé inférieur) d’Angleterre et de Belgique. Bulletin de l’Institut Royal des Sciences naturelles de Belgique, Série Sciences de la Terre, 69:77–96.
Taverne, L. 2006. Révision d’ Ichthyotringa africana, poisson marin (Teleostei, Aulopiformes) du Crétacé supérier de la Mésogée eurafricaine. Considérations sur les relations phylogénétiques du genre Ichthyotringa . Belgian Journal of Zoology, 136:31–41.
Tsujita, C.J. and Westermann, G.E.G. 1998. Ammonoid habitats and habits in the Western Interior Seaway: a case study from the Cretaceous Bearpaw Formation of southern Alberta, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology, 144:135–160.
https://doi.org/10.1016/S0031-0182(98)00090-X
Verma, O. and Khosla, A. 2019. Developments in the stratigraphy of the Deccan Volcanic Province, peninsular India. Comptes Rendus Geoscience, 351:461–476.
https://doi.org/10.1016/j.crte.2019.10.002
Webb, E.J. 1984. Sedimentology of the Upper Cretaceous Coffeesands in northcentral Mississippi. Gulf Coast Association of Geological Societies, Transactions, 34:277–286.
Weiler, W. 1972. Fisch-Otolithen aus der unteren Kreide Niedersachsens. Bericht der Naturhistorischen Gesellschaft zu Hannover, 116:71–79.
Whitehead, P.J.P. 1981. Albulidae. In Fischer, W., Bianchi, G., and Scott, W.B. (eds.), FAO species identification sheets for fishery purposes. Eastern Central Atlantic fishing areas 34 and Part of 47, Volume I. Food and Agriculture Organization of the United Nations, Rome.
Whitehead, P.J.P. 1990. Pterothrissidae, p. 125. In Quéro, J.C., Hureau, J.C., Karrer, C., Post, A., and Saldanha, L. (eds.), Check-list of the fishes of the eastern tropical Atlantic. CLOFETA. Unesco, Paris.
Wilson, M.V.H. and Chalifa, Y. 1989. Fossil marine actinopterygian fishes from the Kaskapau Formation (Upper Cretaceous: Turonian) near Watino, Alberta. Canadian Journal of Earth Sciences, 26:2604–2620.
https://doi.org/10.1139/e89-222
Woods, M.A., Mortimore, R.N., and Wood, C.J. 2012. The chalk of Suffolk, p. 105–31. In Dixon, R. (ed.), A Celebration of Suffolk Geology: GeoSuffolk 10th Anniversary Volume, GeoSuffolk, Ipswich.
Wollemann, A. 1904. Die Fauna des mittleren Gaults von Algermissen. Jahrbuch der Preußischen Geologischen Landesanstalt und Bergakademie, 24:22–42.
Yokoi, T. 1998. Cretaceous Otolith from Sakasa-river, Northern Hokkaido. Chigakukenkyu, 47:101–104. [In Japanese]
