INTRODUCTION
Sub-decadal climate variability in the Pacific Ocean is affected by interdependent variability in the Aleutian Low (AL), North Pacific High (NPH), the Jet Stream, and the equatorial El Niño/La Niña cycle. Superimposed on this variability are less well-understood, regionally modified, decadal and centennial scale cycles that appear to arise from global-scale teleconnections
(Mann et al. 1995;
Ware and Thomson
2000). There is also a growing body of empirical evidence that there is correlation between climate records and solar and cosmic ray activity and their proxy indicators (e.g., geomagnetic field activity, sunspot numbers, and
10Be, 36Cl, 14C abundances) suggesting that extraterrestrial phenomena may be responsible for at least a portion of climate variability (e.g.,
Bond et al. 1997;
Bond et al. 2001;
Kromer et al. 2001;
Neff et al. 2001;
Sharma 2002;
Hu et al. 2003;
Usoskin et al.
2003).
 The instrument record is barely adequate for analyzing characteristics of decadal-scale Holocene climate cycles, including cycles linked to climate change events such as the well-publicized collapse of west coast North American commercial fisheries
(Hare et al.
1999). Because long-term trends in regional and global temperatures are modified by low-frequency events that are difficult to resolve with relatively short historical records, a more complete understanding of the dynamics of natural climate variability as well as medium and long-term climate oscillations can only be achieved by turning to the geological record.
The instrument record is barely adequate for analyzing characteristics of decadal-scale Holocene climate cycles, including cycles linked to climate change events such as the well-publicized collapse of west coast North American commercial fisheries
(Hare et al.
1999). Because long-term trends in regional and global temperatures are modified by low-frequency events that are difficult to resolve with relatively short historical records, a more complete understanding of the dynamics of natural climate variability as well as medium and long-term climate oscillations can only be achieved by turning to the geological record.
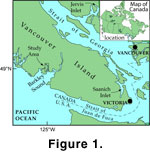
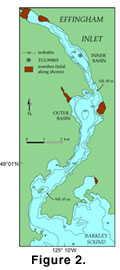 Changes in productivity in Effingham inlet, southwest coast of Vancouver Island, British Columbia, Canada
(Figure 1, Figure
2) are represented in the sediments by variation in the relative thickness of the seasonally deposited laminations of diatom-ooze and terrigenous material. These variations, in turn, can be linked to changing ocean climate conditions
(McQuoid and Hobson
2001).
Changes in productivity in Effingham inlet, southwest coast of Vancouver Island, British Columbia, Canada
(Figure 1, Figure
2) are represented in the sediments by variation in the relative thickness of the seasonally deposited laminations of diatom-ooze and terrigenous material. These variations, in turn, can be linked to changing ocean climate conditions
(McQuoid and Hobson
2001).
The clockwise circulating NPH dominates the NE Pacific region in the summer resulting in cool and dry northerly winds, while the anticlockwise circulating AL dominates during winter, generating warm and moist southerly winds
(Thomson 1981;
Beamish 1993;
Mantua et al. 1997;
Ware 1991; Ware and Thomson 1991,
2000;
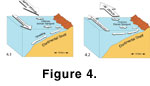
Figure 3). Seasonally dependent upwelling (caused by NPH wind-driven, Ekman transport of surface waters away from the Vancouver Island continental margin, usually between May and August at present) is the principal mechanism providing nutrient supply to the coastal ocean off southwest British Columbia
(Thomson and Gower 1998;
Figure 4). 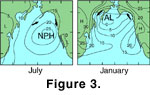 Over time, changing climatic conditions impact the AL and NPH, leading to changes in surface wind stress, regional circulation, mixed layer depth, and stratification. These changes may be at least partially the result of extraterrestrial climate forcing, because there is a link between the 11-year Schwabe sunspot cycle and the centers of action of both the AL and NPH
(Christoforou and Hameed 1997;
Hameed and Lee
2003). The influence of extraterrestrial climate forcing through centennial to millenial-scale cycles are magnified by as much as a factor of four
(Carslaw et al.
2002).
Over time, changing climatic conditions impact the AL and NPH, leading to changes in surface wind stress, regional circulation, mixed layer depth, and stratification. These changes may be at least partially the result of extraterrestrial climate forcing, because there is a link between the 11-year Schwabe sunspot cycle and the centers of action of both the AL and NPH
(Christoforou and Hameed 1997;
Hameed and Lee
2003). The influence of extraterrestrial climate forcing through centennial to millenial-scale cycles are magnified by as much as a factor of four
(Carslaw et al.
2002).
 Changing climatic conditions in the NE Pacific in turn, modify the surface layer of the ocean where phytoplankton growth is concentrated
(Brodeur and Ware
1992). This drives cyclical changes in primary production, timing of blooms and phytoplankton assemblages
(Ware and Thomson
2000). Because pelagic fish respond to food availability, these changes cascade through the various trophic levels eventually impacting commercially important pelagic fish populations
(Beamish and Boillon
1993).
Changing climatic conditions in the NE Pacific in turn, modify the surface layer of the ocean where phytoplankton growth is concentrated
(Brodeur and Ware
1992). This drives cyclical changes in primary production, timing of blooms and phytoplankton assemblages
(Ware and Thomson
2000). Because pelagic fish respond to food availability, these changes cascade through the various trophic levels eventually impacting commercially important pelagic fish populations
(Beamish and Boillon
1993).
The purpose of this research is:
- To determine the implications of the observed fish productivity cycles and the temporal variability in fish-scale abundances of two particularly abundant
taxa, Pacific Herring and Northern Anchovy, in Core TUL99B-03 from the inner basin of Effingham Inlet
(Figure 2). Both Pacific Herring and
Northern Anchovy undergo significant periodic population fluctuations. However, because of the brief modern fishing records, the frequency and amplitude of longer term changes are largely unknown
(Holmgren-Urba and Baumgartner 1993;
Schweigert 1995;
Tunnicliffe et al.
2001).
- Demonstrate the relevance of paleontological analysis in ecological studies. Paleontology is advantageous in ecological analysis because the duration of most biology-based ecological studies are too short to assess the long-term impact of either anthropogenic or natural perturbations on biological communities
(Martin 1991,
1995;
Martin et al. 1999;
Jackson 1992;
Aronson and Precht 1997;
Kowalewski et al.
1999).
Our analysis is also intended to bring the continuous wavelet transform time series analysis methodology (described in more detail
below) to the attention of the paleontological community, which has had little exposure to this useful way of dealing with non-stationary cycles in biostratigraphic records
(Prokoph et al. 2000,
2001).
Pacific Herring and Northern Anchovy Ecology
 Pacific
Herring (Clupea harengus pallasi) is a cool water species
(Figure 5). When annual water temperatures are below average, the typically plentiful herring stock around Effingham Inlet increases in abundance
(Ware 1991;
Schweigert
1995). In contrast, the Northern Anchovy
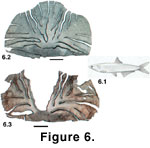
(Figure 6) is typically at the northern limit of its range in British Columbia. Anchovy are more productive in this region when upwelling is suppressed and sea surface temperature (SST) is elevated
(Richardson
1981). In the absence of major biotic factors such as predation and overfishing recruitment, mortality rates and abundance of anchovy and herring populations around Effingham Inlet should vary in opposition to one another as a function of changes in upwelling and SST conditions. Alongshore advection may also play a role because upwelling-favorable southerly winds give rise to southward mean currents over the slope and outer shelf, while downwelling-favorable northward winds cause northward mean currents in the region.
Pacific
Herring (Clupea harengus pallasi) is a cool water species
(Figure 5). When annual water temperatures are below average, the typically plentiful herring stock around Effingham Inlet increases in abundance
(Ware 1991;
Schweigert
1995). In contrast, the Northern Anchovy
(Figure 6) is typically at the northern limit of its range in British Columbia. Anchovy are more productive in this region when upwelling is suppressed and sea surface temperature (SST) is elevated
(Richardson
1981). In the absence of major biotic factors such as predation and overfishing recruitment, mortality rates and abundance of anchovy and herring populations around Effingham Inlet should vary in opposition to one another as a function of changes in upwelling and SST conditions. Alongshore advection may also play a role because upwelling-favorable southerly winds give rise to southward mean currents over the slope and outer shelf, while downwelling-favorable northward winds cause northward mean currents in the region.
Pacific Herring
 BC herring populations are spring spawners (March/April) and tend to spawn at temperatures around 7-8ºC. Recruitment to the West Coast of Vancouver Island herring population tends to be much lower when ocean temperatures are warm. Accordingly, the abundance of this population declines and tends to be well below average during warm periods. In contrast, average recruitment to this population is twice as high during cool periods, so the population becomes more abundant
(Ware 1991;
Ware
and McFarland 1995).
BC herring populations are spring spawners (March/April) and tend to spawn at temperatures around 7-8ºC. Recruitment to the West Coast of Vancouver Island herring population tends to be much lower when ocean temperatures are warm. Accordingly, the abundance of this population declines and tends to be well below average during warm periods. In contrast, average recruitment to this population is twice as high during cool periods, so the population becomes more abundant
(Ware 1991;
Ware
and McFarland 1995).
Northern Anchovy
Northern Anchovy are distributed from Magdalena Bay, Baja California to the Queen Charlotte Islands, BC. This species is at the northern end of its range in BC. Anchovies appear to prefer water temperatures between 14.5 and 18.5ºC
(Hart
1973).
In the NE Pacific, three populations of anchovy are recognized: the northern (Oregon/Washington/BC), Central (California/Mexico) and Southern (Mexico).
A large component of the northern population of anchovy spawn in waters around the Columbia River plume. Spawning conditions differ between the northern and central populations of anchovy. "Off Oregon, spawning occurs from mid-June to mid-August, when current flow to the south is at a maximum,
water temperatures are reaching maximum levels for the year, coastal upwelling is at a maximum, and day length is at or near maximum duration. Off California, peak spawning occurs from January through April when southward current flow is minimal, water temperatures are reaching minimal levels for the year, upwelling is minimal, and day length is at minimum duration"
(Richardson
1981).
Hart (1973) states that in BC the life history of anchovy "is not well known and occurrence is sporadic." He also notes that anchovies are "not available abundantly or consistently enough in British Columbia to be used commercially" except during a brief period of abundance in the Strait of Georgia in the early 1940s.
An article in the Pacific Biological Station Progress Report (1945) 63:41 states that "the
Northern Anchovy is found in large schools along the British Columbia coast during the spring and early summer." In BC, anchovy are summer spawners. They require water temperatures above 13ºC for successful reproduction. These conditions only occur in a few places in BC and only in the summer (July and August;
Hart
1973). Although water temperatures have been above normal around Barkley Sound since the mid 1970s, the abundance of the anchovy population in the Barkley Sound area has been relatively small. Therefore, other important factors besides warm summer temperatures are required for the successful reproduction and expansion of the local anchovy population around Effingham Inlet (see below). It is unknown whether the Barkley Sound anchovy population is self-sustaining (and somewhat genetically distinct), or whether it is sustained by a periodic influx of migrants from the much larger Columbia River population to the south. Genetic samples would have to be taken to determine if the Barkley Sound population is relatively isolated, or if it mixes with, and is therefore genetically indistinguishable, from the Columbia River population. Because the fish-scale record suggests that Effingham Inlet always contains some anchovies, if this small, local population disappears periodically it must be recolonized fairly quickly by migrants from nearby populations.
An analysis of ocean climate time series off the SW coast of Vancouver Island for the period 1930-45, when anchovies were abundant in BC, indicates that during this period, upwelling favorable winds in the summer (mean JJA wind stress = 0.151
dyn/cm2) were anomalously low (-0.7 standard deviations below the long-term mean JJA wind stress = 0.223; SD=0.106;
Hsieh et al.
1995). During the same period, the summer (JJA) SST was average (a mixture of warm and cool years; DFO Lighthouse data). Therefore, the data clearly indicate that summer upwelling (and, coincidentally, southward alongshore coastal currents) was anomalously weak when historical sources reported that anchovy were abundant in southern BC. Consistent with this interpretation, anchovy have not been abundant in BC since the early 1940s, and coastal upwelling in the summer has not been as weak as it was during that period
(Hart 1973;
Therriault et al. 2002).
A northward shift in the sea surface temperature isotherms during the spawning season (particularly during warm El Niño years such as 1982/83), causes the distribution of the central anchovy population to shift northward
(Fiedler et al.
1986). By analogy, the distribution of the northern population may also shift northward into BC during anomalously warm summers.
Richardson's (1981) paper provides evidence that warm water temperatures are necessary for the successful reproduction of the northern population of anchovy. However, warm summer temperatures alone are not a sufficient condition.
Research summarized by Ware (1991) and
Ware and
McFarland (1995) indicates that the West Coast of Vancouver Island herring population increases in abundance during cool periods and declines during warm periods. Since the herring and anchovy populations around Effingham Inlet should vary in opposition to one another, this
implies that ocean conditions were probably warmer than normal when anchovy were dominant. Historical data also indicate that anchovies were more abundant in BC when summer upwelling was anomalously low. It is also important to appreciate that changes in the abundance of fish scales in a single sediment core could also reflect changes in migration/distribution patterns. Extensive research confirms that the herring around Effingham Inlet do not migrate large distances. They migrate seasonally to offshore feeding banks in the summer, and return inshore in the late Fall to over-wintering areas near the spawning grounds
(Taylor
1964). Consequently, a large increase in herring scales in Effingham Inlet most likely reflects an increase in the productivity of the West Coast of Vancouver Island herring population.
In contrast, under some conditions, it is possible that anchovy could migrate northward from the main spawning area off the Columbia River to the West Coast of Vancouver Island in the summer to spawn and to forage. Consequently, a high abundance of anchovy scales in Effingham Inlet could reflect either: 1) an increase in the abundance of the local population, or an increase in the abundance and northward expansion of Columbia River anchovies (note that in both cases, the increase in abundance arises from an increase in anchovy productivity either locally or to the south of Barkley Sound); or 2) a change in the summer distribution pattern of Columbia River
Anchovy. For example, when summer upwelling is extremely weak, anchovy off Washington may migrate northward to southern BC in search of food.
Scale Preservation
Engraulis mordax mordax (Northern Anchovy)
Scales of the Northern Anchovy preserve easily in Quaternary sediments
(Figure 6). They are robust and have a high degree of mechanical flexibility. They are, however, subject to erosion and fracturing along points of weakness
(Figure 6). The degree of abrasion in the sediments controls the degree of fragmentation. Wings (posterior fields) are often disarticulated, as is the central lobe in the anterior field. In some cases, all three lobes and wing are disarticulated, leaving only the central focal area. Care must be taken when determining counts so that a single fragmented scale is not counted several times. The scales are generally amber in color. Some geochemical conditions can "bleach" them white and cause them to become extremely brittle
(Patterson et al.
2002).
Clupea harengus pallasi (Pacific Herring)
Clupeid scales are, comparative to other fish species, thick and robust and therefore preserve easily in Quaternary/Holocene sediments
(Figure 5). Because of their robustness, herring scales tend to preserve whole, sometimes even with the posterior field (wing) attached
(Patterson et al.
2002). That generality aside,
Clupea scales tend to rip and break at fracture areas. The leading edges of the anterior field and the wing clips are also often detached. As a result it can often be quite difficult to distinguish
Clupea and Sardinops fragments. However, Clupea scales (including the wings) are generally white and quite mechanically flexible, and to some extent easily ripped, while
Sardinops scales are often amber and brittle.
The Data
 Our dataset comprised measurements of laminae thicknesses and laminae gray-scale values extracted from x-ray images of annually deposited sediments, as well as fish-scale abundances
(Figure 7). We applied two time series analysis tools (CWT and SA) to our dataset in order to overcome limitations inherent to each
(Emery and Thomson
2001). CWT has advantages over SA in that it determines the frequency content of time series as a function of time. If temporal changes occur in frequency and phase of time series (i.e., time series are non-stationary), we can recognize and quantify them using CWT analysis. On the other hand, SA provides us with estimates of the "power" (signal variance per unit frequency band) as a function of frequency, as well as estimates of the cross-spectral power between two time series
(Davis
1986). The combined result is a more robust and quantitative periodicity analysis.
Our dataset comprised measurements of laminae thicknesses and laminae gray-scale values extracted from x-ray images of annually deposited sediments, as well as fish-scale abundances
(Figure 7). We applied two time series analysis tools (CWT and SA) to our dataset in order to overcome limitations inherent to each
(Emery and Thomson
2001). CWT has advantages over SA in that it determines the frequency content of time series as a function of time. If temporal changes occur in frequency and phase of time series (i.e., time series are non-stationary), we can recognize and quantify them using CWT analysis. On the other hand, SA provides us with estimates of the "power" (signal variance per unit frequency band) as a function of frequency, as well as estimates of the cross-spectral power between two time series
(Davis
1986). The combined result is a more robust and quantitative periodicity analysis.
