RESULTS
Size and Shape among Caryocorbula,
Bothrocorbula, and Hexacorbula
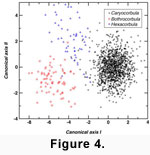 A CVA of uniform and non-uniform shape components reveals that each genus of the
Caryocorbula + (Bothrocorbula + Hexacorbula) clade is morphologically distinct both in shape
(Wilks' lambda = 0.002, p<0.0001; 98.9% of specimens correctly assigned to genus;
Figure 4) and size (Figure
5). Qualitatively, these genera are most readily distinguished by sculptural differences and by the presence of a lunule in
Bothrocorbula (Figure 2.8, Figure 2.10). CVA results highlight more subtle but consistent differences related to the shape of valve outlines among genera (see also
Figure 2). At one extreme on CVA Axis 1,
Bothrocorbula has a slightly narrowed keel and a very broadly rounded anterior margin. In addition, the dorsal ends of the adductor muscle scars, especially the anterior, are positioned more toward the center of the valve.
A CVA of uniform and non-uniform shape components reveals that each genus of the
Caryocorbula + (Bothrocorbula + Hexacorbula) clade is morphologically distinct both in shape
(Wilks' lambda = 0.002, p<0.0001; 98.9% of specimens correctly assigned to genus;
Figure 4) and size (Figure
5). Qualitatively, these genera are most readily distinguished by sculptural differences and by the presence of a lunule in
Bothrocorbula (Figure 2.8, Figure 2.10). CVA results highlight more subtle but consistent differences related to the shape of valve outlines among genera (see also
Figure 2). At one extreme on CVA Axis 1,
Bothrocorbula has a slightly narrowed keel and a very broadly rounded anterior margin. In addition, the dorsal ends of the adductor muscle scars, especially the anterior, are positioned more toward the center of the valve. Bothrocorbula and Hexacorbula separate primarily on the second CVA axis.
Hexacorbula has a narrower and more dorsally positioned rostral area accompanied by posterior expansion of the shell behind the
umbo. Caryocorbula separates from the other genera primarily on CVA Axis 1 and differs most notably in having a much broader rostral area.
Bothrocorbula and Hexacorbula separate primarily on the second CVA axis.
Hexacorbula has a narrower and more dorsally positioned rostral area accompanied by posterior expansion of the shell behind the
umbo. Caryocorbula separates from the other genera primarily on CVA Axis 1 and differs most notably in having a much broader rostral area.
Relative warp analysis of the three genera highlights interspecific allometry in
Caryocorbula that is not observed in either Bothrocorbula or Hexacorbula (illustrated in
Figure 6 using internal landmarks; Note: because we used landmark techniques, the relationship of size and shape is not due to the use of shape variables containing size information, a pervasive problem for distance metrics).
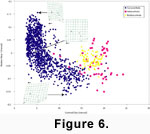 The CVA axes do not record this allometry because
non-allometric features related to the dorso-ventral breadth of the rostral area (broad in
Caryocorbula, narrower in Hexacorbula and Bothrocorbula)
separate the genera in the analysis. In contrast, the first Relative Warp axis describes shape deformation related to valve elongation and rostrum position. Valves with high positive scores on Relative Warp Axis 1 are, relative to the standard reference form, compressed
anteroposteriorly, have adductor muscle scars and ventral point of the pallial sinus (Landmark D on
Figure 3) that are directed inward and upward, and have a ventrally displaced rostrum
(Figure 6). Valves with high negative values on Relative Warp Axis 1 are dorsoventrally compressed with a strongly upturned rostrum, have posterior adductor muscle scars that are displaced inward and downward, and the ventral point of the pallial sinus positioned upward and inward.
The CVA axes do not record this allometry because
non-allometric features related to the dorso-ventral breadth of the rostral area (broad in
Caryocorbula, narrower in Hexacorbula and Bothrocorbula)
separate the genera in the analysis. In contrast, the first Relative Warp axis describes shape deformation related to valve elongation and rostrum position. Valves with high positive scores on Relative Warp Axis 1 are, relative to the standard reference form, compressed
anteroposteriorly, have adductor muscle scars and ventral point of the pallial sinus (Landmark D on
Figure 3) that are directed inward and upward, and have a ventrally displaced rostrum
(Figure 6). Valves with high negative values on Relative Warp Axis 1 are dorsoventrally compressed with a strongly upturned rostrum, have posterior adductor muscle scars that are displaced inward and downward, and the ventral point of the pallial sinus positioned upward and inward.
Juvenile corbulids tend to be extremely thin valved until they approach adult size (Goodwin et al. 2003). As a result, preservation of juveniles should be extremely rare, and the specimens used for all genera in these analyses probably represent valves of adults. Therefore, the absence of observed allometry in
Bothrocorbula and Hexacorbula may reflect the absence of small species in these genera (i.e., allometry not obvious over relatively narrow size ranges; see
Figure 5). The lack of small species, however, indicates that size is an important morphologic feature in Bothrocorbula and
Hexacorbula in spite of the absence of observed interspecific allometry in their valves.
Interspecific Allometry in Caryocorbula
 Neogene Caryocorbula from the Caribbean expresses a strong interspecific pattern of allometry
(Figure 6, Figure
7). The CVA Axis 1 illustrated in Figure
7, like the Relative Warp Axis in Figure
6, describes valve elongation and rostral position, although the axes are reversed. Therefore, small species have relatively low CVA Axis 1 scores and tend to be shortened anteroposteriorly with a ventrally positioned rostrum. Larger specimens are progressively more elongate through dorsoventral shortening and have a more dorsally positioned to upturned rostrum
(Figure 7).
Neogene Caryocorbula from the Caribbean expresses a strong interspecific pattern of allometry
(Figure 6, Figure
7). The CVA Axis 1 illustrated in Figure
7, like the Relative Warp Axis in Figure
6, describes valve elongation and rostral position, although the axes are reversed. Therefore, small species have relatively low CVA Axis 1 scores and tend to be shortened anteroposteriorly with a ventrally positioned rostrum. Larger specimens are progressively more elongate through dorsoventral shortening and have a more dorsally positioned to upturned rostrum
(Figure 7).
 This allometric trend persists throughout the Neogene, as illustrated when specimens from six broad time slices are plotted separately or when allometric curves for time slices are compared
(Figure 8, Figure 9; Note: time intervals in these figures were not analyzed separately but are displayed separately to highlight temporal patterns). In
Figure 8, allometric trajectories (fit with logarithmic equations) for each time slice generally follow the overall allometric trend (black line on
Figure 8), although the overall trend can be offset (i.e., y-intercepts differ) from one time interval to another. A notable exception is the "middle" Pliocene interval (purple line on
Figure 8) where size is somewhat truncated, and where
Caryocorbula sp. C, which has a morphology that does not fall along the general allometric trend, is found (orange diamond in
Figure 7 and Figure 9;
further discussed below and in the Discussion section).
This allometric trend persists throughout the Neogene, as illustrated when specimens from six broad time slices are plotted separately or when allometric curves for time slices are compared
(Figure 8, Figure 9; Note: time intervals in these figures were not analyzed separately but are displayed separately to highlight temporal patterns). In
Figure 8, allometric trajectories (fit with logarithmic equations) for each time slice generally follow the overall allometric trend (black line on
Figure 8), although the overall trend can be offset (i.e., y-intercepts differ) from one time interval to another. A notable exception is the "middle" Pliocene interval (purple line on
Figure 8) where size is somewhat truncated, and where
Caryocorbula sp. C, which has a morphology that does not fall along the general allometric trend, is found (orange diamond in
Figure 7 and Figure 9;
further discussed below and in the Discussion section).
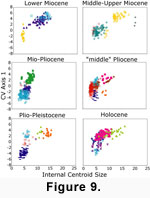 The constancy of this allometric trend is not dependent on a few abundant long-lived species
(Figure 9). Instead, different species of a given size from different localities or times tend to have very similar shapes. For instance,
C. sericea (from the upper Pliocene Bowden, Moin, and Escudo de Veraguas formations of Jamaica, Costa Rica, and Panama, respectively) and
C. lavalleana (from the lower Pliocene Gurabo and Mao formations of the Dominican Republic and Holocene collections from Cuba and Venezuela) anchor the bottom of the curve from the Mio-Pliocene to Holocene, but re-occupy morphospace vacated by another species,
Caryocorbula sp. A, from the upper lower to lower middle Miocene Cantaure Formation of Venezuela
(Figure 9). In addition to species turnover, the allometric pattern in
Caryocorbula persists in spite of uneven geographic and environmental sampling over time (see
Table 2; Appendix).
The constancy of this allometric trend is not dependent on a few abundant long-lived species
(Figure 9). Instead, different species of a given size from different localities or times tend to have very similar shapes. For instance,
C. sericea (from the upper Pliocene Bowden, Moin, and Escudo de Veraguas formations of Jamaica, Costa Rica, and Panama, respectively) and
C. lavalleana (from the lower Pliocene Gurabo and Mao formations of the Dominican Republic and Holocene collections from Cuba and Venezuela) anchor the bottom of the curve from the Mio-Pliocene to Holocene, but re-occupy morphospace vacated by another species,
Caryocorbula sp. A, from the upper lower to lower middle Miocene Cantaure Formation of Venezuela
(Figure 9). In addition to species turnover, the allometric pattern in
Caryocorbula persists in spite of uneven geographic and environmental sampling over time (see
Table 2; Appendix).
Three time intervals show some variance from the overall allometric pattern. First, the middle-upper Miocene interval is missing small caryocorbulids with high negative values on CVA Axis 1 but includes the largest
Caryocorbula from the Caribbean Neogene. (This is also the interval in which corbulid generic diversity peaks in the Caribbean
[Anderson 2001;
Anderson and Roopnarine
2003].) Second, in the Mio-Pliocene, the allometric curve is truncated dramatically, without an apparent change in curve shape
(Figure 8, Figure
9). This extinction of large-bodied
Caryocorbula species is part of an extinction of large species of all corbulid genera present in the Caribbean. These extinctions have been attributed to a decrease in nutrient availability and primary productivity in the Caribbean that commenced at this time
(Anderson
2001). Third, the "middle" Pliocene, as well as the middle to upper Miocene, each contain a handful of individuals from species (Caryocorbula sp. B and
Caryocorbula sp. C) that are offset from the overall allometric trend (Figure
7, Figure 9).
There are few similar studies to which we can compare our results, making it difficult to infer how common or rare such interspecific allometric trends are. Although studies of allometry and scaling are common, most studies track allometry within species, and of those that track allometry among species most studies focus on covariation in the size (i.e., length, mass, volume) of two traits (i.e., those of the
Huxley-Jolicouer School). Very few studies have documented the relationship of size and shape among species (i.e., the
Gould-Mosimann School). For the "Huxleyan" data sets, interspecific allometry is relatively common, and observed in vertebrates in general
(Jerison
1973) and in terrestrial mammals (Christiansen
2002), primates (Majoral et al.
1997), deer (Gould
1974), and horses (Radinsky
1984). For the few "Gouldian" data sets (including two studies using the geometric morphometric techniques we employ;
Penin and Berge 2001,
Rosenberg
2002), a range of allometric trends among species are documented including differing allometric slopes for species (e.g.,
Rosenberg
2002), similar trends that offset along the size axis (similar to the pattern we see for species pooled by time interval,
Figure 8; e.g., Emlen
1996), or a combination of differing trends and trend offsets (e.g., Kawano
2000).
