COMPARISON WITH JURASSIC SPECIES ATTACHED TO SPONGES
In this section, modern taxa examined above are compared with Jurassic sponge facies foraminifera to find possible "equivalents." By "equivalent" we mean either having a close taxonomic relationship, a similarity in external morphology or a similarity in the habitat they colonize. Jurassic sponge species that are absent or rare in the Recent will be discussed at the end of the section.
Arenaceous/Calcareous Ratio
Both in the Jurassic and in the Recent, the percentage of arenaceous taxa is higher among the sponge fragment dwellers than in the surrounding mud. The modern reefs, however, contain a larger proportion of calcareous taxa than the Jurassic reefs. After subtracting the always-loose taxa (Table 3), the calcareous individuals in our material are nearly as numerous as the arenaceous (810 against 880). Calcareous forms are abundant in modern reefs despite the fact that 1) the waters off British Columbia are certainly colder than the subtropical northern Tethys and 2) in the carbonate-laying environment in which the sponge reefs developed, postmortem dissolution of CaCO3 is likely to have been slower than on the modern British Columbia shelf. It is possible that silicification of Jurassic calcareous foraminifera was poor—this would have affected the quantitative results of etched sponge studies though it is impossible to say how much. Bias in the representative value of
Table 3 should not have affected the arenaceous/calcareous ratio. An obvious factor is that some of the very common modern taxa, Lobatula and Islandiella, did not exist in the Jurassic and may have since moved into niches formerly occupied by arenaceous forms.
Arenaceous Species
By its growth form and wall characteristics, Ammobaculinus recurvus resembles the widespread Jurassic genus Haplophragmium. Ammobaculinus differs from this last taxon only by its aperture. Haplophragmium-like forms are uncommon in the Recent: the only other genus is Acupeina
Brönniman and Zaninetti
(1984) (multiple aperture) from shallow, brackish tropical waters. Among the authors that extracted silicified foraminifera from Jurassic sponges, none reported Haplophragmium.
Crithionina is not reported from Jurassic reefs. This may be due to the tendency of some of its species to disaggregate postmortem.
The phenomenon of trapped foraminifera in the Jurassic is observed mostly among the genera Thurammina, Tolypammina and Subbdelloidina. Silicified foraminifera extracted from the limestone by etching show marks left on the tests by the presence of the spicules. Such specimens are described by German authors as eingeschnürt, or "laced in" (Seibold and Seibold 1960a,
1960b;
Schmalzriedt 1991;
Munk 1994).
 Subbdelloidina haeusleri
Frentzen (1944) is a close Jurassic equivalent of Placopsilina spongiphila. Both grow attached to rods of the meshwork. The specimens illustrated by
Frentzen (1944) and by
Seibold and Seibold (1960a) differ from P. spongiphila by their generally larger diameter, more depressed sutures and primarily, their tendency to branch (see
Appendix). The illustrations seem to indicate that they are "pseudoattached" (sensu
Hofker 1972, quoted by
Gooday and Haynes 1983) whereas P. spongiphila is generally "attached." We re-photographed the specimens of
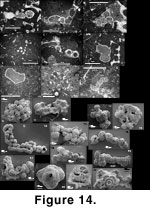
Schmalzriedt (1991) and illustrate them on
Figure 14.15-14.19.
Subbdelloidina haeusleri
Frentzen (1944) is a close Jurassic equivalent of Placopsilina spongiphila. Both grow attached to rods of the meshwork. The specimens illustrated by
Frentzen (1944) and by
Seibold and Seibold (1960a) differ from P. spongiphila by their generally larger diameter, more depressed sutures and primarily, their tendency to branch (see
Appendix). The illustrations seem to indicate that they are "pseudoattached" (sensu
Hofker 1972, quoted by
Gooday and Haynes 1983) whereas P. spongiphila is generally "attached." We re-photographed the specimens of
Schmalzriedt (1991) and illustrate them on
Figure 14.15-14.19.
Schmalzriedt (1991) synonymizes Subbdelloidina with Placopsilina. He reports two species of Placopsilina but since he sees all intermediates between both, we believe they should be considered as morphotypes of the same species. Whether this species should be included under Placopsilina or Subbdelloidina should wait for a review of Placopsilina. In the meantime, we will go on using the name Subbdelloidina haeusleri.
Schmalzriedt's smallest specimens of Subbdelloidina have about the same diameter as P. spongiphila (~100 µm) and fill only a part of the space inside the meshwork; more commonly, they are 200 µm or more and fill most of the mesh space, depending on the sponge species they were colonizing. As a result, Subbdelloidina is contorted and shows obvious traces of the presence of the sponge meshwork. As P. spongiphila, it does not show any tendency to lift its test from the substrate, which suggests it fed on the substrate. The S. haeusleri illustrated by
Munk (1994) are fairly large, often with a tangled (knäuelig) initial part, and may be attached to other foraminifera. The initial part of P. spongiphila is often a sort of tangle but it is wound around an intersection of the meshwork, something not usually seen in S. haeusleri.
In Jurassic sponge reefs, the genus Placopsilina (not Subbdelloidina) occurs as large specimens encrusting the exterior of sponge fragments. This form and habit is absent from our material with the possible exception of the single above-mentioned specimen from the Strait of Georgia (Figure 4.16-4.17).
If Telammina-like forms ever inhabited Jurassic sponge fragments, it is likely that attempts at extracting them by etching would have destroyed the delicate stolons (assuming these were preserved) and left only indeterminable agglutinated balls.
Valvulina lobata, a species very close to ?Tritaxis fusca was described from the Jurassic sponge reefs by
Seibold and Seibold (1960a) and later transferred to Tritaxis by
Oesterle (1968). Tritaxis lobata is common and is one of the few attached species inside Jurassic sponge fragments.
Wagenplast (1972) reports it as Valvulina sp. from the material etched out of the sponges, but does not find it in the surrounding marl (Schwammmergel).
Schmalzriedt (1991) reports it (as Tritaxis) as abundant inside the sponges themselves, as single specimens elsewhere in the reef, and as absent off-reef. Tritaxis lobata occasionally shows a large attachment cyst with one or more apertures (Schmalzriedt 1991;
Munk 1994) which we do not find in our modern specimens. Such a structure would be typical of Trochamminella.
Gaillard (1983) reports it (as Valvulina) as exclusive to the sponge facies (equivalent to the reef-facies of Schmalzriedt) where it is fairly common.
Munk (1994) illustrates the apertural side of T. lobata and shows a specimen attached to a Thurammina, itself contorted, having grown inside the meshwork. The main difference between T. lobata and our modern specimens is that the periphery of the former is lobate and irregular; moreover, T. lobata has 2½ chambers in the last whorl, like T. fusca (Seibold and Seibold 1960a). These differences are minor: these are two closely related taxa living attached in and on sponge meshwork, 150 Ma apart.
Trochamminids have been widely reported from Jurassic etched sponge faunas. They show approximately the same range of thickness-to-diameter ratio, sharpness of periphery, chamber inflatedness, and number of chambers in the last whorl as the morphotypes from modern sponges. The Jurassic species illustrated in the literature show no trace of attachment or entrapment as we see in our modern specimens.
Calcareous Species
Chilostomella oolina, Globobulimina auriculata and Nonionella digitata had not appeared yet in the Jurassic, but other streamlined forms were present, Guttulina, Eoguttulina, Dentalina and Nodosaria. They have been found both in the reef facies and off-reef but only
Schmalzriedt (1991) reported them from etched sponges, as "occasional."
Cassidulinidae appeared only in the Tertiary and are thus absent in Jurassic sponge reefs. Other mid-sized calcareous species are reported but those from inside the sponges are few in numbers; this could be explained by postmortem dissolution as is the case with Islandiella in our material. The specimens illustrated (in particular by
Schmalzriedt 1991) appear poorly preserved, but this is more likely the result of incomplete silicification than of etching of the CaCO3.
The genus Lobatula did not exist in the Jurassic. Calcareous attached forms were probably Trocholina and Paalzowella (Patellina? in
Schmalzriedt 1991), but their test wall is quite different from that of Lobatula and the preservation/dissolution may have differed too. These genera are occasional (Schmalzriedt 1991), or locally frequent (Seibold and Seibold 1960a) in the etched sponges, and are often totally absent. The clinging (i.e., epifaunal) genus Spirillina is very abundant around Jurassic sponges but less so inside, and its presence there could be accidental (loose fauna). The suspension feeder niche in the dead Jurassic sponges appears mostly occupied by Tritaxis and Tolypammina.
Kazmierczak (1973) and
Gaillard (1983) have discussed the question of whether foraminifera on Jurassic reefs are commensal organisms or postmortem settlers. Our specimens of Lobatula, even when they suggest the former presence of soft parts, are firmly attached to the meshwork and nothing in their position suggests that they were inside a canal of the living sponge benefiting of the food particles carried by the currents produced by the sponge, as
Kazmierczak (1973) believed for Tolypammina.
Ramulina siphonifera must be compared with the pictures of
Schmalzriedt (1991, plate 1, figure 9 re-illustrated in the present
Figure 14.13) which show a Jurassic Thurammina similarly punched with holes, and with the original illustrations of Thurammina canaliculata (Haeusler 1883). Thurammina is the only Jurassic taxon to fit our concept of "impaled."
Some species of Ramulina and Bullopora are associated with Upper Jurassic sponge reefs. Ramulina fusiformis
Khan (1950), and Ramulina spandeli
Paalzow (1917) have been found in the sponge reef facies and the bank facies but not in the sponges themselves (Seibold and Seibold 1960a;
Gaillard 1983). These do not have the contorted aspect of R. siphonifera nor its very thin siphon and are quite different. Bullopora rostrata
Quenstedt (1857) grows attached on the exterior of sponges; its closest equivalent in modern sponge reefs would be Lobatula sp. rather than R. siphonifera. By contrast, Bullopora tuberculata (Sollas
1877), another attached form, is quite close to R. siphonifera. Both grow intertwined with the meshwork. The modern form is rarely found anywhere else, whereas the Jurassic species may also be present away from the sponge reefs, at various water depths (Septfontaine 1977;
Gaillard 1983;
Schmid 1996). Both have an irregular shape consisting of a succession of constrictions and widenings (see illustrations in
Gaillard 1983 and
Schmid 1996). In R. siphonifera, this results mostly from the presence of the meshwork through which it creeps and does not represent a true succession of chambers. No connection between successive chambers has been found in R. siphonifera, contrary to B. tuberculata (Figure 14.2-14.3). Both have remarkably similar conical spines—which are more representative of Ramulina than of Bullopora—though we have not been able to check whether the spines of R. siphonifera have a central canal like those of B. tuberculata. Our modern specimens have a rather thin wall, less calcification being normal in cool waters with predominantly clastic sedimentation, compared with the subtropical Jurassic sea in which carbonate sediments were deposited. The most obvious difference is the thin apertural tube; such a structure has not been reported from the Jurassic Bullopora. The overall exterior outlook of B. tuberculata can be only reconstructed from thin sections as it has never been extracted from the sponge fragments; however, something as characteristic as a thin protruding tube would undoubtedly have been reported by some of the authors working on Jurassic faunas.
Bullopora tuberculata is common in thin sections but absent in the silicified residues. It is possible that lenticulinids became silicified while B. tuberculata did not, but it is not clear why it would be so as both have a similar wall structure and composition. It is possible both are silicified but that researchers working with thin sections have sampled different levels than investigators of silicified foraminifera because of differences in working methods and in sampling goals, and that the latter sampled essentially beds without Bullopora.
We would not pretend that R. siphonifera and B. tuberculata are synonyms; there are clear morphologic differences. We would not even argue that the first descends directly from the second. Despite the obvious taxonomic changes since the Jurassic, it is remarkable that, after 150 Ma, dead sponges continue to offer the same habitat and that different organisms adapt to it in the same way, generating similar morphologies that are found nowhere else.
In the Jurassic sponge reef facies, Bullopora rostrata may seem to be the closest equivalent to aff. Oolina caudigera. Both belong to the Nodosariaceae, are subspherical and attach to dead sponges. However, B. rostrata is plurilocular and limited to the outside of sponge fragments (never found in the etched assemblage) and it is a true attached form, like Lobatula. On the contrary, the attachment of aff. Oolina caudigera is accidental; it is more impaled than attached, and on this point resembles many other forms in this material that are not normally known to attach (Gaudryina, Dorothia, etc.). Its closest Jurassic equivalent would thus be the genus Thurammina.
Jurassic "Sponge Foraminifera" Rare or Absent in Modern Reefs
A certain number of taxa commonly reported from sponge reefs by Jurassic authors (Table 6) are not discussed above. They are either totally absent or rare in the modern sponge fragments, or they are important enough in the Jurassic to deserve a separate discussion. The species discussed below were selected on the basis of our experience in the Jurassic as well as a review of the literature.
Vinelloidea crussolensis
Canu (1913) (Figure 14.4-14.5). Often reported as Nubeculinella bigoti
Cushman (1930) (for taxonomy see
Voigt 1973;
Loeblich and Tappan 1988), this is probably the most abundant foraminiferal species to attach to Jurassic sponges. It is found on the outer surface of sponges along with abundant serpulid worms and comparatively rare Placopsilina. It is not observed intertwined in the meshwork. It may encrust hard surfaces on the Jurassic seafloor far from any sponge reef, as long as sedimentation rate is low (Gaillard 1983).
Members of the subfamily Nubeculinellinae are widespread in the Recent but their distribution is limited to warm temperate to tropical waters. Queen Charlotte Sound, with its temperatures not exceeding 7°C, is not the kind of habitat where one would expect Nubeculinellinae.
Vinelloidea is closely associated with stromatolitic layers that encrust the upward facing side of sponges; nothing else but a few pelecypods can be found in these crusts.
Hiller (1964),
Hiller and Kull (1967), and later
Gaillard (1983) suggested a possible symbiotic relationship within the crust, which they consider to be built by photosynthetic algae, and Vinelloidea. In more recent literature (Neuweiler and Reitner 1993;
Leinfelder et al. 2002) these crusts are interpreted as microbially induced carbonate precipitations (automicrites) for which light is not necessary. Even though stromatolitic crusts still exist today, for example in Shark Bay, Australia, the deep water, stromatolite-reinforced sponge mound biotope disappeared gradually after the end of the Jurassic. This may have been the cause for the extinction of V. crussolensis, which was adapted to this very specific ecological niche.
As a group, miliolids are rare in our modern sponge reefs. Of the 17,000 foraminiferal specimens identified in the sponge reefs, only 36 were miliolids. Of these, only three were found in the sponge fragments, one of which was a deep-water Pyrgo. This cold and deep environment is probably not the kind that will attract a rich and varied miliolid assemblage, except for species that prefer it (ex.: Pyrgo vespertilio
[Schlumberger, 1891b]) and
for a
few cosmopolitan forms (ex.: Cyclogyra). However, some miliolids can be found in the Arctic, and cool temperatures alone cannot explain their quasi-absence on the British Columbia shelf.
The assemblages obtained by etching are not reported to contain Vinelloidea or any other miliolid test, as if they did not become silicified postmortem. It is possible also that the Vinelloidea-rich stromatolitic layers encrusting the sponges were not sampled by authors who studied etching residues.
"Tubiphytes" morronensis
Crescenti (1969). This form, reported by
Schmid (1996) as attached (even impaled: Schmid's figure 116) to siliceous sponges, has never been reported in strata younger than Mesozoic. Its wall is characterized by a thick, porcellaneous, "micropeloidal" outer layer probably resulting from the action of algal symbionts. Although
Schmid (1996) speculates that it could have lived at depths in excess of 70 m in the tropical Jurassic sea, it is improbable that light penetration on the modern British Columbia shelf could allow algae to develop at 200 m or even at much shallower depths. Thus it is not surprising to find no light-dependent microencrusters
in our modern sponge fragment fauna.
Thurammina sp. (Figure 14.6-14.8). Thurammina sp. is another common species in Jurassic sponge reefs. Thurammina papillata
Brady
(1879) and variants are the only Jurassic sponge-reef foraminifera to be impaled (illustrations in
Schmalzriedt 1991) with rare exceptions (above-mentioned "Tubiphytes"). Thurammina was recognized as pierced with holes by
Hauesler (1883), but he thought these holes were canals.
Seibold and Seibold 1960a,
1960b) recognized these holes as the trace of sponge spicules. This is due to the test being so large as to engulf a few cells of the meshwork and even to bulge beyond that, the same way Ramulina siphonifera does in the Recent. Note that the originally siliceous sponge meshwork was transformed to CaCO3 early in diagenesis, and that sample etching yielded only silicified foraminiferal tests bearing scars or holes left by the former presence of spicules.
Thurammina papillata lives in the Recent but we found none in the sponge reefs, despite looking carefully for it. This niche seems to be occupied now by many different species, R. siphonifera, K. bradyi and Lobatula spp. being the most common. Ramulina siphonifera is the only one for which we positively know that it wraps every silica rod it engulfs with its test wall. Some other species possibly do, but determining this will require careful dissection.
Tolypammina vagans (Brady
1879) (Figure 14.9). This is one of the most common species in the Jurassic sponge reefs (Table 6). It is also one of the rare Jurassic foraminiferal species still present in modern oceans, but it is absent from sponge reefs.
Seibold and Seibold (1960a) believe that their Tolypammina is not the same as the modern T. vagans and therefore report Tolypammina spp.
Wagenplast (1972) does not determine his foraminifera beyond genus level.
Kazmierczak (1973) reports T. vagans from inside sponges where he thinks they were collecting food carried by the currents circulating through the sponges' pores.
This is the second main trapped Jurassic species. It grows by winding its way through the meshwork. The Jurassic sponge meshwork commonly reported by the above-mentioned authors has often smaller cells than our modern sponges, and given the rather large diameter of its tubes, T. vagans is often tightly trapped within the meshwork (illustrations in
Schmalzriedt 1991 and the "Hyperammina contorta" of
Haeusler 1890). The few slender ?Tolypammina sp. we report in our modern material have a quite different growth habit from the Jurassic T. vagans due to their smaller diameter (40-50 µm). Placopsilina spongiphila creeps through the meshwork, but it is attached to the meshwork, not trapped. The fact its aperture never lifts from the substrate may be an indication that it collects food lying around on the meshwork and is not a suspension feeder as T. vagans. We occasionally observe large agglutinated tubes, probably Polychaete worms, meandering through the lattice in a way that reminds us of the Jurassic T. vagans. However, polychaete worms are probably too different from foraminifera to be considered as ecological equivalents.
?Nodophthalmidium sp.
Schmid (1996) reports this Ophthalmidium-related form from Upper Jurassic sponge reefs (Table 6). We find it quite commonly, in thin sections, in the Jurassic sponge facies of the Swabian Alb. It is not far from "Tubiphytes" nor from Nodobacularia or Vinelloidea, but it is not a form that is known to be attached. We did not find any related form on the modern British Columbia shelf, but this is not surprising as conditions there are not favourable for miliolids except for a few taxa.
Textularia spp., Bigenerina spp., Ammobaculites spp., Gaudryinella sp. and Reophax spp. These taxa are common in the Jurassic sponge facies and reported by most of the authors of
Table 6. They are all arenaceous, elongate, uniserial or biserial (at least in the later parts), and their length is in the 400-1000 µm range approximately. Gaudryina uvigeriniformis
Seibold and Seibold 1960a should be included in this group. This constitutes a group that is crudely comparable to the "moderately large arenaceous" taxa of our modern material: Gaudryina, Martinottiella, Karreriella, Dorothia, Ammobaculinus and Reophax. The modern specimens are commonly trapped or impaled. By contrast, the Jurassic specimens illustrated in the literature do not show evidence of having been laced-in (eingeschnürt). They may have been slender enough to fit within the meshwork without pressing against it, provided the mesh is large enough. This however corresponds to the definition of "loose fauna." A large collection of Jurassic sponge foraminifera would have to be examined looking specifically for scars due to the meshwork.
Thomasinella? pauperata (Haeusler) emend.
Oesterle (1968). This taxon of uncertain affinity was first described as Reophax pauperata by
Haeusler (1885). It is reported as "Thomasinella" pauperata by
Schmalzriedt (1991). It resembles Placopsilina spongiphila by its growth habit but it is nearly three times the diameter; also, it is vertically flattened and laterally "keeled." It was probably not creeping inside the lattice but instead grew attached to the surface of sponge fragments. No trace, scar or hole due to the presence of the lattice is mentioned nor shown. This taxon is known mostly from fragments, which makes any inference about its mode of life difficult. We found no comparable form in our modern material.
Cribrostomoides spp. + Recurvoides spp. We would place many of the specimens illustrated by
Oesterle (1968) and
Schmalzriedt (1991) under the name Haplophragmoides in the genus Cribrostomoides (because of the aperture's position) or even Recurvoides (because of bent coiling plane:
Figure 6.22). These genera are present in small numbers in our material and in larger numbers in the Jurassic etched sponges. The species are not the same as in the Recent. Our most common form is Cribrostomoides jeffreysi, and it is not quite like any of the reported Jurassic species. Many of Jurassic forms are small and could be part of the Jurassic "loose" fraction though
Schmalzriedt (1991) reports none from the "normal" (i.e., bedded or bank) facies.
Spirillina spp. and Lenticulina spp. These are common open shelf genera in the Upper Jurassic and are reported as part of the sponge fauna (Table 6). Spirillina in particular has been reported by all workers as being particularly abundant in the immediate surroundings of sponge reefs. Collectively, these forms occur in percentages that are comparable to those of the most common species in the mud retained by modern sponges: Epistominella vitrea, Bolivina decussata, Eponides pusillus, Seabrookia earlandi and Angulogerina spp. Whenever they are small enough, they may be found "loose" inside the meshwork (Table 3). It is impossible to say whether they crept in by themselves or were brought in accidentally, but their presence is not surprising considering their abundance in the mud. Contrary to what happens with modern samples, the sieving process cannot be held responsible for the introduction of Spirillina and the smaller lenticulinids into the meshwork of Jurassic sponges. These two genera could be part of the Jurassic "loose" fauna. The larger lenticulinids could be like some of the moderately large modern specimens whose situation cannot be definitely assigned to the "loose" or "trapped" category. We think the report of "globigerinids" in the Jurassic etched sponge facies (Wagenplast 1972) can be explained in the same way; we also find occasional planktonics in our modern sponge fragments.
