CORRELATION AND BIOSTRATIGRAPHY OF THE KHAN FORMATION
WITH CORRELATIVE PERI-GONDWANA DEPOSITS
Twenty-one samples were collected from four sections at 1 m intervals, and 352 oriented thin-sections with fusulinids were prepared. The recovered assemblage of fusulinids consists of six genera and 24 species. Six genera of smaller foraminifera were found as well (Table 1). The most abundant taxa in the assemblage are diverse Eoparafusulina, and Perigondwania; the Neodutkevitchia are much less diverse. Schubertellides (Eoschubertella and Grovesella) and staffellids are very rare but present in the assemblage. Smaller foraminifera in the studied material are very rare.
Eoparafusulina, Perigondwania, and Neodutkevitchia that are distributed within Peri-Gondwana appear there as the earliest, relatively warm water marine assemblage above cool water carbonates. They are associated with brachiopods and bryozoans and cold/cool-water siliciclastic deposits including glacial diamictites (Angiolini et al. 2006;
Leven 1993,
1997;
Ueno 2003;
Shi and Grunt 2000). This assemblage represents the transition between non-fusulinid cold-water communities and typical warm-water fusulinid communities of a tropical area. Accordingly, a stressful environment can be expected in this transition that is marginal for fusulinids. These conclusions are based on the following observations. First, the diversity of the assemblage is relatively low relative to the assemblages in the Tethys of a comparible age (e.g., Afghanistan, North and South China). Second, an abnormally large number of the specimens (>5% of the total population) have a double proloculi. A similar phenomenon was observed in the published data from Central Pamir (Leven 1993). From the 102 figured specimens of Eoparafusulina, Perigondwania and Neodutkevitchia in this paper, six specimens have a double proloculi (Leven 1993, Plate 1, figure 7; Plate 2, figure 5; Plate 5, figure 9; Plate 7, figure 16; Plate 11, figures 6 and 14). The unusually large number of specimens with a double proloculi might indicate a stressful environment that is marginal for these surviving fusulinids. In normal warm-water conditions the number of specimens with double proloculi never exceeds 0.1% of the population.
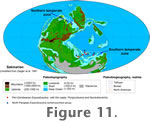 Two assemblages can be recognized in the Kalmard area. One is Eoparafusulina-dominated and found in Gachal and Madbeiki sections. The second, a Perigondwania-Neodutkevitchia-dominated assemblage is found in the Halvan and Rahdar sections. The Eoparafusulina-dominated assemblage consists of eight species of Eoparafusulina, seven of which are new, and one species of Neodutkevitchia (Table 1). One previously known species is E. pamirensis (Leven 1993). It has been found in the Kalmard area and was originally described from bed 6 of Dangikalon Fm in Central Pamir (Leven 1993), where it appears in an assemblage with several other Eoparafuslina, numerous Perigondwania, less diverse Neodutkevitchia and perhaps rare Pseudoendothyra, Schubertella and Pseudoreichelina (Leven 1993). Unfortunately, no documentation about species distribution in each section/sample nor ecological or environmental information were provided in this publication. It is hard to judge whether Eoparafusulina in Central Pamir appear together with Perigondwania and Neodutkevitchia or whether these two assemblages occur separately as in the Kalmard area (Figure 11).
Two assemblages can be recognized in the Kalmard area. One is Eoparafusulina-dominated and found in Gachal and Madbeiki sections. The second, a Perigondwania-Neodutkevitchia-dominated assemblage is found in the Halvan and Rahdar sections. The Eoparafusulina-dominated assemblage consists of eight species of Eoparafusulina, seven of which are new, and one species of Neodutkevitchia (Table 1). One previously known species is E. pamirensis (Leven 1993). It has been found in the Kalmard area and was originally described from bed 6 of Dangikalon Fm in Central Pamir (Leven 1993), where it appears in an assemblage with several other Eoparafuslina, numerous Perigondwania, less diverse Neodutkevitchia and perhaps rare Pseudoendothyra, Schubertella and Pseudoreichelina (Leven 1993). Unfortunately, no documentation about species distribution in each section/sample nor ecological or environmental information were provided in this publication. It is hard to judge whether Eoparafusulina in Central Pamir appear together with Perigondwania and Neodutkevitchia or whether these two assemblages occur separately as in the Kalmard area (Figure 11).
Two new species of Eoparafusulina (E. grozdilovae and E. stevensi) that are found in the Kalmard area also occur in bed 6 of Dangikalon Fm in Central Pamir and were designated there as Eoparafusulina tschernyschewi tschernyschewi and Eoparafusulina tschernyschewi memoranda (Leven 1993). The latter two species were originally described from late Asselian-early Sakmarian of Timan (Schellwien 1909;
Grozdilova and Lebedeva 1961). It is plausible that neither population of E. grozdilovae nor E. stevensi from Central Pamir or from the Kalmard areas belong to species from the Arctic for several reasons. First, they differ from Arctic Eoparafusulina in morphology. The thickness of septa in all Eoparafusulina from the Arctic is as thick as a spirotheca (Grozdilova and Lebedeva 1961), whereas septa in Eoparafusulina from Central Pamir and the Kalmard areas are twice or more thinner than spirotheca. It seems that thin septa are characteristic for all other Eoparafusulina in the Peri-Gondwana regions (Premoli-Silva 1965;
Leven 1993,
1997;
Gaetani et al. 1995;
Ueno 2003). Second, Eoparafusulina from the Arctic, Central Pamir and Kalmard areas possess a different stratigraphic range: late Asselian-early Sakmarian vs. latest Sakmarian-early Artinskian. Third, these two groups of Eoparafusulina are distributed within two separate antitropical temperate high latitude belts on opposite sides of the tropical belt (Figure 11). That is, Eoparafusulina from the Arctic occur in the northern antitropical belt, whereas Eoparafusulina from the Peri-Gondwana regions occur in the southern antitropical belt. In early Sakmarian, the northern transitional temperate zone of the Arctic became completely isolated from either Tethys and/or Peri-Gondwana (Davydov and Leven 2003) and consequently Arctic Eoparafusulina had no path for migration into Peri-Gondwana during the latest Sakmarian-early Artinskian. An independent origin of Eoparafusulina in Peri-Gondwana is the most probable scenario. However, no direct evidence is available.
Eoparafusulina is widely distributed in the Peri-Gondwana and known from the Baoshan and Tengchong Blocks in Yunnan (Ueno 2003; Shi Yukun personal commun., 2006), the Rutog area, South Tibet (Nie and Song 1983a), East Hindu Kush (Gaetani et al. 1995), Karakorum (Premoli Silva 1965), South and Central Afghanistan (Leven 1997), central Pamir (Leven 1993) and Oman (Angiolini et al. 2006). Leven also reported several species of Eoparafusulina in Central Pamir that were originally described in Alaska and the Shasta Lake Area of North America (Petocz 1971;
Skinner and Wilde 1965). Although they possess remarkable similarity to some specimens from Central Pamir (Leven 1993), these are most probably different and new species. For example, Eoparafusulina depressa
Skinner and Wilde 1965 and E. laudoni
Skinner and Wilde 1965 from Shasta Lake Area possess much better developed cuniculi in 2-3 outer volutions whereas cuniculi in Eoparafusulina from Peri-Gondwana are always less well developed and can be barely observed only in final volution.
The Perigondwania-Neodutkevitchia dominated assemblage is distributed within Peri-Gondwana approximately in the same regions as the Eoparafusulina-dominated assemblage. In the Kalmard area the diversity of this assemblage is relatively poor. Six species of Perigondwania are found there and are also known in South Tibet (Nie and Song 1983b), East Hindu Kush (Gaetani et al. 1995), Karakorum (Premoli Silva 1965), South and Central Afghanistan (Leven 1997), central Pamir (Leven 1993)
and Oman (Angiolini et al. 2006). The two species of Perigondwania from the Kalmard area (P.rahdariensis and P. aghanabati) as well as all Neodutkevitchia are new.