|
|
|
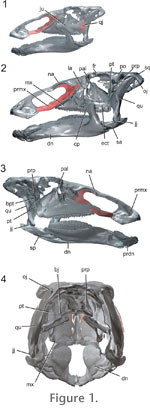
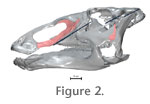
INTRODUCTIONReconstructing feeding behavior in extinct animals is fundamental to gaining insight into their ecology and evolution. Previous investigations into dinosaur feeding have utilized multiple approaches including, soft-tissue reconstruction (Ostrom 1961; Haas 1963; Galton 1984; Norman 1984; Molnar 1991) and study of toothwear patterns (e.g., Norman and Weishampel 1985; Weishampel and Norman 1989; Fiorillo 1998; Barrett 2000). Additionally, consideration of joint surfaces have served to help reconstruct kinematics of the feeding system, and more recently finite-element modeling has been used to investigate how the head skeleton may resist strains associated with feeding (e.g., Ostrom 1964; Weishampel 1984; Upchurch and Barrett 2000; Rayfield et al. 2001; Rayfield 2004; Therrien et al. 2006). A major challenge in reconstructing dinosaurian feeding behavior relates to our incomplete understanding of soft tissues (e.g., musculature, ligaments) (Witmer 1995). Even if the gross morphology of the soft tissues is properly reconstructed, it is still difficult to infer how the soft tissues would have interacted with the head skeleton to produce feeding movements (Lauder 1995). The challenge appears most acute for lineages where the head skeleton is highly derived, as seen in hadrosaur dinosaurs. Nonetheless, the architecture of the head skeleton does offer numerous data that can be used to identify constraints in mobility, an important first step in any kinematic reconstruction. Within Dinosauria, feeding behavior of hadrosaurs has garnered particular attention. Hadrosaurs (Ornithopoda, Iguanodontia, Hadrosauridae) are a globally successful group of large-bodied, Late Cretaceous herbivores (Ostrom 1961; Norman and Weishampel 1985). A possible key to their success is a specialized feeding system that includes a transverse chewing stroke (Norman and Weishampel 1985). Transverse chewing in combination with other specializations such as a robust occluding dentition, may have allowed hadrosaurs to process tough foods more effectively than their less specialized herbivorous counterparts. Among tetrapods, transverse chewing appears to have arisen within only two lineages: mammals (Weijs 1994) and hadrosaurs. In mammals, transverse chewing is achieved through mandibular movements, whereas in hadrosaurs it has been hypothesized to involve a unique, complicated set of intracranial movements (i.e., pleurokinesis) (Weishampel 1984; Norman and Weishampel 1985). Despite many previous studies into the hadrosaur feeding apparatus (Ostrom 1961; Heaton 1972; Norman 1984; Weishampel 1984) elucidating the mechanisms of cranial kinesis has been difficult in part because of conflicting interpretations of the soft tissues (Ostrom 1961; Weishampel 1984), but also because of the complex geometry associated with the proposed cranial kinesis. This study is the first to investigate hadrosaur feeding by combining three-dimensionally scanned, cranial elements with animation techniques. The advantage of this 3-D animation approach is that it allows us to take into account the geometry of each cranial element and investigate how the architecture of the skull and shapes of the intracranial joints constrains feeding movements. Pleurokinesis and the evolution of the hadrosaur feeding apparatus
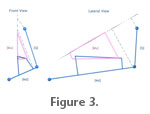
The pleurokinetic hypothesis of iguanodontid chewing assumes bilateral occlusion (i.e., isognathy) and a unique system of mobile intracranial joints (Norman and Weishampel 1985).
This study represents the first step toward developing a new animation model that will be used to investigate and test chewing hypotheses in Edmontosaurus. Here we describe how the animation model was created from scanned fossil elements recovered from a single individual, and then mobilized using inverse kinematics and a wiring system. We finish with a discussion of the preliminary results and an outline of future directions. Notably, an additional objective of this study was to produce an animation, which could illustrate the pleurokinetic hypothesis of hadrosaur chewing for museum visitors. The model described herein is currently on exhibit at the Canadian Museum of Nature, Talisman Fossil Gallery in Ottawa.
|
|