|
|
|
DISCUSSIONThere are general differences in tooth and jaw movement among the groups examined here, with both the occlusal profile and the degree of lateral movement of the condyle showing that greater lateral excursion of the jaw and teeth occur in Groups 3 and 4 compared to Groups 1 and 2. For none of the groups is the occlusal path of the protoconid linear, particularly towards the beginning of the occlusal phase after the teeth make contact, during which the carnassial blades are occluding. Therefore, the occlusal vector changes during the slow-closing phase. The jaw movement simulated here for these species indicates that the jaw movement is associated with the number of teeth in the tooth row and the approximate complexity of the teeth. The more simple tooth rows (Groups 1 and 2) are those that show more vertical movement, and those with more lateral movement have a greater number of teeth and cusps (Groups 3 and 4). Groups 1 and 2 have fewer teeth (only one or two teeth in each jaw), and these teeth have only two or three main cusps, whereas groups 3 and 4 have a greater number of teeth (two or three in each jaw), which tend to have a larger number of cusps on them (see Figure 3, Figure 4, Figure 5, and Figure 6). These general differences in complexity are also reflected in quantitative measurements of dental complexity such as orientation patch count (OPC; Evans et al. 2007): species in Groups 1 and 2 have an average OPC of 91 and 56.25 for the upper and lower tooth rows, respectively, while the corresponding values for Groups 3 and 4 are 163 and 115.25. The domestic cat is the species that is the most applicable comparison with the current study, as it appears to be the only carnivoran in which a detailed study of jaw movement and electromyography during mastication has been carried out (Gorniak and Gans 1980). It is difficult to compare the reconstructions of jaw movement directly to the domestic cat as there is insufficient resolution at the end of the closing stroke using cinematographic methods to determine the fine-scale path of the tooth. However, the sequences generally show a lingual movement of the jaw at the very end of the sequence, which is consistent with these reconstructions. During the slow-closing phase of these occlusal reconstructions, tooth-tooth contact between the upper and lower tooth rows occurs at the same approximate positions as the major attrition facets on the teeth. It is difficult to be precise about the areas of contact between the teeth in the reconstruction as very high resolution scans could not be used due to limitations of computer power. This supports the contention that observed jaw movements are capable of producing the observed wear facets, and therefore, that an examination of these wear facets will reveal a significant amount regarding occlusal relations. Therefore, the incorporation of the three-dimensional position and orientation of both the overall tooth shapes and the attrition facets will bring a much greater understanding to this endeavour.
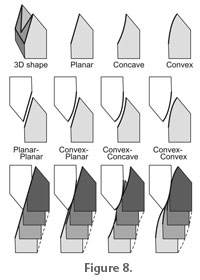
For the planar-planar and convex-convex shapes, continual contact will be maintained between the upper and lower blades through the occlusal stroke, giving effective cutting between the blades. However, for the other two combinations illustrated (convex-planar and convex-concave), the blades will be separated at the beginning and/or end of the stroke, depending on the exact shape of the facets. This will presumably lead to inefficient blade-on-blade cutting by these two combinations. Attrition facets indicate tooth-tooth contact and therefore that there is no relief or space between the trailing surfaces of the blades at that point. In newly-erupted carnivoran teeth, attrition facets are absent. This generally means that there is relief behind the blades, which is functionally desirable as it will reduce jamming or friction between the blades (Evans and Sanson 2003; Evans 2005). We found that the lower molars in the Group 1 species had a higher facet curvature towards the tips of the cusps. This could aid in initial alignment of the carnassials at the beginning of the stroke. If the attrition facets at the tip of the cusp tended to be flat, very precise alignment would be required to ensure that the opposing blades came into contact close to the tip of the cusps, i.e., the area of the lower tooth that the upper tooth could contact, which we could term the 'target area', is quite small. However, a higher curvature towards the tips of the cusps increases the 'target area' for one blade to align with the other at the beginning of the stroke. This will give a greater tolerance for initial alignment. After that, alignment can be controlled by the opposing facet shapes. The models of carnassial tooth shape constructed by Evans and Sanson (2006) have a linear occlusal path, the blades are two-dimensional in that they lie in a plane, and relief behind the blades is generated by curving the relief surface out of the plane in which the blade lies. Substantial tooth-tooth wear on the relief surface of such a tooth (and little wear on the rake or leading surface) would generate a large attrition facet (see figure 7 in Evans 2005). This will be disadvantageous to the function of the tooth, allowing food to be trapped between the relief surfaces, and requiring more force to maintain contact between occluding blades than when there is relief space behind the cutting edge. Many of the specimens examined in this study have large attrition facets (particularly Acinonyx and Crocuta), implying that there are large areas of tooth-tooth contact, potentially reducing the effectiveness of the teeth. However, the attrition facets for these two species are convex (non-planar), and as the tooth-tooth contact proceeds during the virtual chew, only a small amount of the attrition facet is in contact between the teeth at any one time. This means that the instantaneous facet contact is much lower than the actual facet size, like the convex-convex combination in Figure 8. Relief behind the cutting edge is, therefore, maintained due to the reciprocally curved geometry of the attrition facets and the occlusal movement of the teeth. Relief would be present in the convex-planar and convex-concave combinations when the blades are in contact, but on average will be greatest for the convex-convex combination. The facts that the attrition facets for some of these species are not planar, and the occlusal path is not linear, are consistent with one another: non-planar (non-extruded curve) facets would indicate a non-linear occlusal path and vice versa. The obvious difference between the simplified attrition facets in Figure 8 and those of the real teeth are that there is rotation around a condyle rather than a direct translation up the page, as shown in the figure. Rotation around a condyle would still give a planar surface for a straight crest or a 'fan-shaped' plane for another shape. The curvature of this fan would depend on the distance from the centre of rotation. The occlusion reconstructions demonstrate that if the only rotation allowed is around the lateral axis, then lateral movement of the jaw within the glenoid fossa is required. For those species that appear to show large degrees of lateral jaw movement in these reconstructions, rotation around a second axis, such as the dorso-ventral or mandibular axes, may also occur. Tooth occlusion could then take place without as much lateral translation of the condyles along the glenoid fossae, and this will affect the resulting facet shape. The degree of fusion of the symphysis and mobility at the CMJ will influence how much rotation in other axes is possible. The shape of the jaw joint surfaces could be used to estimate the position and degree of rotation of the jaw. For species with cylindrical joint surfaces, such as Group 1, it is unlikely that much rotation around the vertical or mandibular axis occurs, but for those where the fit between the joint surfaces is not as tight, this is more likely. A number of limitations in the design of this study should be borne in mind. The tooth and joint positions were scanned from a dry skull, and so post-mortem warping of the skull and mandible means that the positions and orientations of these may have changed. However, the very good fit of most of the teeth during the occlusion stroke suggests that this is not substantial. The necessity of estimating the gap between the jaw joint surfaces to account for the joint capsule also limits the accuracy with which the occlusion in the functioning skull can be simulated. Furthermore, the position and contact between occluding canines has not been modelled in this study. In a number of carnivoran species (as well as in certain primates, suiforms, and representatives of other living and extinct groups), the canine teeth appear to aid guiding the postcanine teeth into occlusion, and limit the lateral excursion of the jaw. Since the main focus of this study was to model how the teeth and jaw move once the postcanine teeth have come into occlusion, this omission should not affect the general findings of the study. This reconstruction of occlusal dynamics in several carnivoran species has established that both the tooth and jaw joint surfaces provide constraints on occlusion, which were Herring's (1993) first two determinants of power stroke direction, and that they are sufficient to give an occlusal path consistent with that described in previous investigations. However, for some species at least, it is apparent that the tooth facets do not form strict inclined planes, but rather surfaces that are slightly curved, which can often be advantageous for tooth function. More detailed analyses of the three-dimensional shape and movement of the mandible and jaw joints in vivo will go further in establishing the fine-scale control and functional consequences of dental occlusion in mammals. |
|