|
|
|
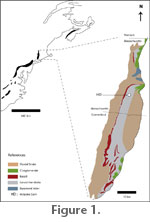
MATERIALS AND METHODSNomenclatural ConsiderationsHistorically, vegetative shoots ascribed to the Araucariaceae have sometimes been described using the generic name Araucarites; however, Zijlstra and Konijnenburg-van Cittert (2000) proposed that this name be restricted to megasporangiate cones and isolated bract-scale complexes. This practice is acceptable when applied to early representatives of the Araucariaceae with uncertain sectional affinities or to more poorly preserved material. However, the name Araucarites should not be applied to material assigned to a particular section, as such a determination must indicate affinity with the genus Araucaria. The generic name Araucaria is used here with reference to the new Holyoke specimen, but it is not formally named as a new species as it is represented by a single, isolated specimen that cannot be definitively diagnosed from several Araucaria section Eutacta bract-scale complexes. Some other comparative taxa are referred to here as Araucarites based on the original descriptions until a thorough revision of the fossil record of the family is available. Locality and Geological Setting
Fossil Preparation and Study Methods
Phylogenetic and Statistical MethodsAlthough the value of fossils in phylogenetic reconstruction is controversial, the time of first appearance of groups in the geological record is widely used in calibrating phylogenies based on molecular evidence. Furthermore, fossils are sometimes used to describe the level of agreement between the temporal sequence of taxa in the fossil record and the order of branching on phylogenic trees. Such congruence studies normally employ one of two approaches; 1) methods that evaluate the number of inconsistencies between the phylogeny and temporal data (Norell and Novaceck 1992; Huelsenbeck 1994), and 2) methods that measure calibrated "ghost lineages" or phylogenetically implied gaps (Sidall 1998; Wills 1999; Pol and Norell 2001) by which the absolute temporal disparity is evaluated (Brochu and Norell 2000). In this paper, the second approach is used to evaluate the congruence between the current phylogenetic hypotheses for the sections of Araucaria and the temporal information from the fossil record of the genus. Specifically, we used the analysis of Setoguchi et al. (1998), which produced one hypothesis of relationships between the sections of Araucaria and that of Gilmore and Hill (1997), which produced two topologies–one perfectly congruent with that of Setoguchi et al. (1998) in terms of the sectional relationships and one different. The Gilmore and Hill (1997) study used fewer Araucaria species, but it included at least one species of each monophyletic section of the genus (sensu Setoguchi et al. 1998). The first appearance datum (FAD) used for each taxon is based on the literature for sections Araucaria, Bunya, and Intermedia (e.g., Stockey 1994; Setoguchi et al. 1998). The FAD for section Eutacta is based on the Holyoke bract-scale complex described here. The Manhattan Stratigraphic Measure (MSM) method originally proposed by Sidall (1998) and later modified by Pol and Norell (2001) designated as MSM* was utilized. In addition, the age uncertainty was considered with the randomization approach for age ranges (Pol and Norell 2006) using the MSM* to calculate the stratigraphic fit for each of 1000 random replicates. This analysis was performed using the phylogenetic analysis software program TNT (Goloboff et al. in press). |
|