|
|
|
INTRODUCTION
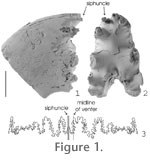
Counterbalance mechanisms were prevalent in straight-shelled Paleozoic cephalopods such as Actinoceratoids and Endoceratoids (Westermann 1998). These groups used calcium carbonate to weight their distal ends in order to orientate their soft bodies away from the dangers of the substrate while improving their hydrodynamic efficiency when moving across the ocean floor. Once these groups became extinct in the Middle to Late Paleozoic, counterbalance mechanisms became obsolete. However, some Jurassic ammonites have asymmetries in the elements of their phragmocone, which raises the question as to whether a counterbalance mechanism was in effect, allowing the animal to remain upright in the water column. Following the end-Triassic mass extinction, Psiloceras formed the root stock for the Jurassic and Cretaceous ammonite radiation (e.g., Guex 2006). In a typical planispirally coiled ammonite, the siphuncle usually ran centrally along the venter of the shell within the plane of bilateral symmetry. However, in many species of Jurassic ammonites, it has long been recognized that the siphuncle can be considerably offset to one side (e.g., Canavari 1882; Swinnerton and Trueman 1917; Spath 1919; Hölder 1956; Hengsbach 1986a, 1986b; Guex 2001). Although some researchers believed it was of no biological importance (e.g., Diener 1912), several biological explanations have been proposed. Spath (1919, p. 116, 118 and references therein) suggested it may be one line of evidence of an attempt to change a bilaterally symmetric swimmer into a crawling benthic organism. In the case of flat ventered shells, Swinnerton and Trueman (1917) hypothesized that asymmetry allowed the siphuncle to take up a stable position along the ventrolateral shoulder. Hengsbach (1979, 1986a, 1986b, 1991, 1996) proposed that an asymmetric siphuncle may be due to the presence of parasites in early ontogeny that caused displacement of the root of the siphuncle. Recent work has highlighted the role of external stress in increasing the rate of gene mutation and recombination as well as in the initiation of asymmetries (Hoffman and Parsons 1991). It seems possible that environmental stress in the earliest Jurassic may have generated asymmetry of the siphuncle in the ammonite root stock (Guex 2001). Subsequently, this character persisted intermittently within the Jurassic part of the ammonoid evolutionary radiation.
|
|