|
METHODS
We developed two types of computer model. The first (model 1), of a single septum and one chamber length of siphuncle, was used to examine the effects of asymmetry within a single chamber of the ammonite's phragmocone. The second type of model is of a complete ammonite, which allowed us to examine the potential effects of phragmocone asymmetries on the entire ammonite animal. In this case, two models of different shell diameter were constructed (models 2 and 3) permitting analysis of the impact of phragmocone asymmetries at different growth stages. All measurements were taken from specimens of B. columbiae. Abbreviations and measurements follow
Smith (1986) and include maximum shell diameter (DMAX) and whorl height (WH). Details of model construction are given below.
Model 1: Components within a Single Phragmocone Chamber
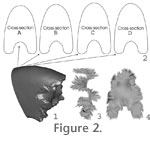 Scanned Fragment. Model 1 was based on an ammonite whorl fragment that had a well exposed septal face (GSC 131683,
Figure 1.1-1.2). A three-dimensional scan of this fragment was created at Memorial University in St. John's, Newfoundland, using a FARO platinum arm with laser line probe. Details of the scanner can be found
on the
Faro website. The scanned data were imported into the three-dimensional CAD modeling program and surface triangulation software, Geomagic. In this program, the scanned model was edited to create a three-dimensional outline of the fossil (Figure 2.1). Digital processing error and surface imperfections such as minor surface cracks caused by weathering were smoothed out manually. The fossil outline was exported to the surface modeling and engineering analysis software Rhinoceros, which is capable of calculating a range of geometric properties including volumes and surface areas of various geometries. Details of the program can be found at
the Rhinoceros website. Several different cross-sections were taken to check for symmetry about the centre line (Figure 2.2). The whorl shape was found to be symmetric within reasonable error (Table 1). Scanned Fragment. Model 1 was based on an ammonite whorl fragment that had a well exposed septal face (GSC 131683,
Figure 1.1-1.2). A three-dimensional scan of this fragment was created at Memorial University in St. John's, Newfoundland, using a FARO platinum arm with laser line probe. Details of the scanner can be found
on the
Faro website. The scanned data were imported into the three-dimensional CAD modeling program and surface triangulation software, Geomagic. In this program, the scanned model was edited to create a three-dimensional outline of the fossil (Figure 2.1). Digital processing error and surface imperfections such as minor surface cracks caused by weathering were smoothed out manually. The fossil outline was exported to the surface modeling and engineering analysis software Rhinoceros, which is capable of calculating a range of geometric properties including volumes and surface areas of various geometries. Details of the program can be found at
the Rhinoceros website. Several different cross-sections were taken to check for symmetry about the centre line (Figure 2.2). The whorl shape was found to be symmetric within reasonable error (Table 1).
Single Septum. Using Geomagic, the septal face was lifted off the fossil scan. Due to erosion, the ventral tips of the saddles were missing from the scan. Thus, the geometry extending from approximately the outer third of the septal
face to the suture line had to be reconstructed. The outermost edges of the scanned septal face were cropped off, and the innermost two-thirds of the septal face were imported into Rhinoceros.
 The septal suture line directly preceding the scanned septal face (Figure 1.1) was used in combination with parts of a suture line from another specimen where necessary, to create a complete suture line (Figure 1.3). This two-dimensional suture line was projected onto the scanned shell and scaled to ensure the peaks and saddles correlated with the septal face and identifiable landmarks on the scan. Approximately 1500 lines tangent to the edge of the septal face were hand-drawn, joining the edge of the septal face to the suture line. Finally, a surface was lofted through these lines resulting in a complete septal face, composed of the scanned central part with modeled edges extending out to the superimposed suture line (Figure 2.3-2.4). The septal suture line directly preceding the scanned septal face (Figure 1.1) was used in combination with parts of a suture line from another specimen where necessary, to create a complete suture line (Figure 1.3). This two-dimensional suture line was projected onto the scanned shell and scaled to ensure the peaks and saddles correlated with the septal face and identifiable landmarks on the scan. Approximately 1500 lines tangent to the edge of the septal face were hand-drawn, joining the edge of the septal face to the suture line. Finally, a surface was lofted through these lines resulting in a complete septal face, composed of the scanned central part with modeled edges extending out to the superimposed suture line (Figure 2.3-2.4).
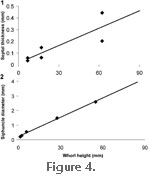
In order to calculate their thickness, thin sections were cut perpendicular to septa (Figure 3.1) at three different shell diameters. It is well recognized that ammonite septa thin from the centre towards the margins of the shell (Westermann 1971). This was accounted for by measuring the thickness at the middle of each septa as well as the thickness of the very outside edge. These thicknesses were then plotted against whorl height, and the average of the two values was used to represent the thickness of the septa at a given whorl height (Figure 4.1).
 The area of the septal face was obtained using a built-in function within Rhinoceros. By multiplying the septal face surface area from the model by the thickness from the graph, a volume was established for the ammonite septum (Table
2). It is well documented that ammonite septa were composed principally of aragonite (e.g.,
Kulicki 1996), which has usually reverted to calcite during diagenesis. Analysis of the measured septa using x-ray diffraction confirmed they had altered to calcite. As volume increases when aragonite is altered to calcite (e.g.,
Liu and Yund 1993), the volumes taken from the model were reduced by 7.6 % before being included in the calculations. This percentage was calculated using the following equations: The area of the septal face was obtained using a built-in function within Rhinoceros. By multiplying the septal face surface area from the model by the thickness from the graph, a volume was established for the ammonite septum (Table
2). It is well documented that ammonite septa were composed principally of aragonite (e.g.,
Kulicki 1996), which has usually reverted to calcite during diagenesis. Analysis of the measured septa using x-ray diffraction confirmed they had altered to calcite. As volume increases when aragonite is altered to calcite (e.g.,
Liu and Yund 1993), the volumes taken from the model were reduced by 7.6 % before being included in the calculations. This percentage was calculated using the following equations:
If density = mass/volume and mass is constant then
Equation (1): volume calcite (A) = 1 g/2.71 g/cm3 = 0.369 cm3
Equation (2): volume aragonite (B) = 1 g/2.93 g/cm3 = 0.341 cm3
Equation (3): volume difference = A - B = 0.028 cm3
Equation (4): total volume change = 0.028 cm3/0.369 cm3 x 100 % = 7.6 %
The density used for the septa was the same as for the shell (2.67 g/cm3,
Hewitt and Westermann 1996) and was used in combination with the adjusted volume to produce a mass of about 4.12 g for the modeled septum (Table 2).
Single Chamber Length of Siphuncle. The length of siphuncle within a single chamber was measured directly from the scanned fragment (25.7 degrees, 33.8 mm,
Figure 1.1). In order to calculate the diameter of the siphuncle, five polished thin sections were cut perpendicular to the siphuncle at different whorl heights (Figure 3.2). In each case, the siphuncle diameter was then measured and plotted against whorl height (Figure 4.2). A best fit line was applied to this graph and measurements were taken from this line to assign the siphuncle diameter to the model. A recent study of Cretaceous phylloceratids found that as ammonites grew, new siphuncular tissue was added in front of the last formed septum (Tanabe et al. 2005). Based on this
finding, the centre of the chamber length of the siphuncle in our model was added such that it intersected the septum. Using the model, the volume was determined by capping each end of the siphuncle and using a built-in function in Rhinoceros (Table 2). The siphuncular tube consists of conchiolin membranes and is very thin (Westermann 1971;
Tanabe et al. 2005). It is primarily filled with blood vessels and surrounding epithelium (Tanabe et al. 2000). For this reason, the density of the siphuncle was taken as 1.055 g/cm3, the same as the density of the soft body of Nautilus (Saunders and Shapiro 1986). The product of density times volume produced a mass of about 0.24 g for the siphuncle on the model (Table 2).
Cameral Sheets. Sheets of organic material have been discovered in the chambers of Triassic, Jurassic and Cretaceous ammonites (Grandjean 1910;
Weitschat and Bandel 1991;
Westermann 1992; and references therein) and may have been present in B. columbiae. Thus, cameral sheets are included mathematically in model 1 as a separate, optional component. Cameral sheets are broadly divided into three groups: horizontal sheets, siphuncular sheets and transverse sheets (Weitschat and Bandel 1991). Horizontal sheets are not included here as they are probably limited to Triassic ammonites with nearly spherical conchs (Westermann 1992). Siphuncular sheets are usually short and straight in Jurassic ammonites and probably functioned to attach the siphuncle to the inner shell wall whereas transverse sheets include all sheets that are subparallel to the septa including some that are suspended from the siphuncular tube (Weitschat and Bandel 1991). As both the latter two types of cameral sheets are associated with the siphuncle, in cases where the siphuncle is offset to one side, their mass would also have been displaced. Cameral sheets probably occupied a total of about 14% of cameral space (Hewitt and Westermann 1996;
Kröger 2002). In model 1, 9.5 % of cameral space is considered to be occupied by cameral sheets (as horizontal sheets were not present). The siphuncular sheets would have been completely offset as they surrounded the siphuncle (3 % of chamber volume was allocated for these). In addition, a portion of the transverse sheets would also have been offset as some of these are suspended from the siphuncle (1.5 % of chamber volume was allocated for these). Thus, in the model, 4.5 % of the volume of the chambers was considered to be taken up by cameral sheets that
were offset whereas the remaining 5 % of chamber volume was considered to be occupied by cameral sheets that were considered central. Total chamber volume was determined directly from the model using the built-in function in Rhinoceros (Table 2). As cameral sheets are believed to have been organic in composition (e.g.,
Erben and Reid 1971;
Weitschat and Bandel 1991 and references therein) and saturated with cameral liquid (Kröger 2002), the density of the cameral sheets is considered to be 1.055 g/cm3, the same as the soft tissue of Nautilus (Saunders and Shapiro 1986). The product of density times 9.5 % of total modeled chamber volume produces a mass of about 3.65 g for the cameral sheets (Table 2).
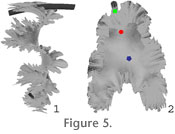 Complete Model. Except for the optional cameral sheets that are only included in calculations, the complete model is shown in
Figure 5. The position of the centroids of the septum and siphuncle were calculated from the model using a built-in function within Rhinoceros (Figure 5.2). In this paper, the centroid of a component is taken as its geometric centre. In the case of the models discussed herein, all the components are considered to have had a uniform density. Thus, the centroid coincides with the centre of mass, and the two terms are used interchangeably throughout the paper. For the cameral sheets, the position of the centroid is taken as the average offset distance of all the sheets weighted according to the percentages discussed above (Figure 5.2). The distances of offset from the centre line are indicated in
Table 2. Complete Model. Except for the optional cameral sheets that are only included in calculations, the complete model is shown in
Figure 5. The position of the centroids of the septum and siphuncle were calculated from the model using a built-in function within Rhinoceros (Figure 5.2). In this paper, the centroid of a component is taken as its geometric centre. In the case of the models discussed herein, all the components are considered to have had a uniform density. Thus, the centroid coincides with the centre of mass, and the two terms are used interchangeably throughout the paper. For the cameral sheets, the position of the centroid is taken as the average offset distance of all the sheets weighted according to the percentages discussed above (Figure 5.2). The distances of offset from the centre line are indicated in
Table 2.
 Model 2: Entire Ammonite Animal at 300 mm Shell Diameter Model 2: Entire Ammonite Animal at 300 mm Shell Diameter
To investigate how these asymmetries affected the entire ammonite animal, a three-dimensional model of a specimen of large shell diameter was constructed (model 2, DMAX = 300 mm). Required data included whorl dimensions as well as septal face surface areas, thickness and number of septal faces, siphuncle diameter, shell thickness and body chamber length. Procedures used to calculate each of these components are addressed below. Direction notations for the model are indicated in
Figure 6.
 Whorl Dimensions. The inner and intermediate portion of the model was created using measurements taken from the specimen of B. columbiae shown in
Figure 7.1 (GSC 127377, DMAX = 197 mm). In addition, a fragment of the outer whorl of a large, incomplete specimen (Figure 7.2, GSC 131683, WH = 132.6 mm) was used to provide information for the model at large shell diameter. This whorl fragment is from the same ammonite as the scanned fragment used as the basis for model 1 but is about 360 degrees larger in size. Whorl Dimensions. The inner and intermediate portion of the model was created using measurements taken from the specimen of B. columbiae shown in
Figure 7.1 (GSC 127377, DMAX = 197 mm). In addition, a fragment of the outer whorl of a large, incomplete specimen (Figure 7.2, GSC 131683, WH = 132.6 mm) was used to provide information for the model at large shell diameter. This whorl fragment is from the same ammonite as the scanned fragment used as the basis for model 1 but is about 360 degrees larger in size.
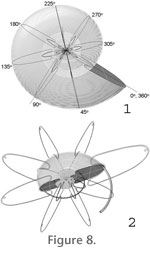 Measurements of the distance of the generating curve from the coiling axis (Raup 1967), whorl height and whorl width were measured at a number of positions and used to provide the model dimensions. These measurements were plotted as a function of angle. The 0 degree position in the model was set as the start of the shell at maximum shell diameter (DMAX = 300 mm), and the outermost 360 degrees of rotation is termed the first whorl. In other words, the angular position increases proceeding towards the umbilicus (Figure 8.1). The original cross-section from the scan was scaled in width and height to match the curve fit through the measured values at 45 degree increments (Figure 8.2). A total of seven whorls for an angle index up to 2520 degrees were drawn; however, given the very small size of the innermost whorl (DMAX = 1.2 mm), only the first six whorls (up to 2160 degrees) were modeled in detail and used for analysis. Measurements of the distance of the generating curve from the coiling axis (Raup 1967), whorl height and whorl width were measured at a number of positions and used to provide the model dimensions. These measurements were plotted as a function of angle. The 0 degree position in the model was set as the start of the shell at maximum shell diameter (DMAX = 300 mm), and the outermost 360 degrees of rotation is termed the first whorl. In other words, the angular position increases proceeding towards the umbilicus (Figure 8.1). The original cross-section from the scan was scaled in width and height to match the curve fit through the measured values at 45 degree increments (Figure 8.2). A total of seven whorls for an angle index up to 2520 degrees were drawn; however, given the very small size of the innermost whorl (DMAX = 1.2 mm), only the first six whorls (up to 2160 degrees) were modeled in detail and used for analysis.
Septal Face Surface Areas. Model 1 provided a septal face surface area of 5098.52 mm3 for a whorl height of 62.3 mm. In order to examine how septal face surface areas changed through ontogeny, two more septal face surface areas were needed. Each of these was calculated using the following equations:
Equation (1): If unknown septal face surface area = A, unknown whorl cross-section surface area = B, septal surface area of scanned face = C and scanned whorl cross-section surface area = D then A = (C x B)/D.
However, A needs to be adjusted to take into account the simplification of the suture line as shell diameter decreases using a simplification factor (equations 2-4).
Equation (2): If suture line length of unknown septal face = E and whorl perimeter length of unknown face = F then E/F = G.
Equation (3): If suture line length of scanned septal face = H and whorl perimeter length of scanned septal face = I then H/I = J.
In order to calculate the suture line length for equations 2 and 3, the suture lines were drawn, flattened and then measured.
Equation (4): The simplification factor is G/J = K.
Equation (5): Therefore, the actual unknown septal face surface area (L) = A x K.
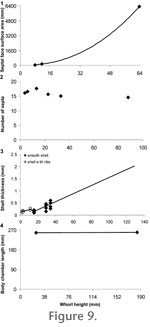 The two calculated surface areas as well as the septal face surface area from the scanned specimen were then plotted against whorl height (Figure 9.1). A curve was fitted through the three values to allow for a septal surface area estimate for each septum. The two calculated surface areas as well as the septal face surface area from the scanned specimen were then plotted against whorl height (Figure 9.1). A curve was fitted through the three values to allow for a septal surface area estimate for each septum.
Septal Thickness and Number. The best fit line for the graph of septal thickness versus whorl height described for model 1 was used to assign a thickness to each septum (Figure 4.1). The number of septa per 360 degrees was counted in a specimen at several ontogenetic stages and then plotted against the whorl height measured at the large end of each revolution (Figure 9.2). For the purposes of model construction, the average number of septa per whorl was taken as 14, and they were placed every 25.7 degrees behind the septum at the back of the body chamber.
Siphuncle Diameter. As in model 1, the best fit line for the graph of siphuncle diameter plotted against whorl height was used to assign diameters to the siphuncle in the model (Figure 4.2).
Although the specimen used to make the shell measurements is quite small (DMAX = 74 mm) relative to the model,
Westermann (1971) showed that shell wall thickness in the Ammonitina is approximately isometric or even positively allometric with shell diameter. Thus, we consider the plot to be reasonably accurate for extrapolation of data at larger shell diameters.
 Shell Thickness. Only one specimen of B. columbiae had an intact shell that was suitable for thickness measurements (Figure 10, GSC 127319). Following the method used by
Kröger (2002), measurements were taken at six different whorl heights. In each case, fragments of shell were removed by hand from five different areas of the whorl; specifically, the umbilical shoulder, lower flank, midflank, upper flank and venter. Each fragment was measured using microcalipers. The average of the five measurements was calculated, and the average thickness value was plotted against whorl height (Figure 9.3). A curve was fitted through the measured values to allow for a shell thickness estimate at each 45 degree increment. There was a slight increase in shell thickness on the innermost whorls due to the presence of ribs in the shell (Figure 9.3). Shell Thickness. Only one specimen of B. columbiae had an intact shell that was suitable for thickness measurements (Figure 10, GSC 127319). Following the method used by
Kröger (2002), measurements were taken at six different whorl heights. In each case, fragments of shell were removed by hand from five different areas of the whorl; specifically, the umbilical shoulder, lower flank, midflank, upper flank and venter. Each fragment was measured using microcalipers. The average of the five measurements was calculated, and the average thickness value was plotted against whorl height (Figure 9.3). A curve was fitted through the measured values to allow for a shell thickness estimate at each 45 degree increment. There was a slight increase in shell thickness on the innermost whorls due to the presence of ribs in the shell (Figure 9.3).
Cameral Sheets. Similar to model 1 described above, cameral sheets were considered as an optional component.
Body Chamber Length. Body chamber length was measured at two different shell diameters. In the first case, a body chamber length of 258 degrees was measured on a small specimen (Figure 9.4, GSC 131684, WH = 25 mm). In the second case, the body chamber length had to be estimated because complete specimens of B. columbiae at large shell diameters are unknown. The large fragment used to create the model has approximating septa and is the final portion of a mature phragmocone (Figure 7.2, GSC 131683, WH = 132.6 mm). This fragment was used in combination with a second incomplete specimen where the final portion of phragmocone is missing but the body chamber is present (Figure 7.3, GSC 127056). By inferring the position of the septate fragment, a body chamber length of about 259 degrees was obtained. Because both the observed and calculated body chamber lengths were very similar, a length of 259 degrees was adopted for the model.
Model Assembly. Once the dimensions of all the components were obtained, the model was assembled using Rhinoceros. Whorls (containing shell and siphuncle outline) of similar absolute angle (i.e., 0, 360, 720, etc. degrees) were fitted together and rotated into position. Surfaces were then lofted onto the shell, body and siphuncle outlines, and ends were put onto the lofted surfaces to make volume calculations possible (Figure 8.2). Finally, the model was divided into whorl or half-whorl portions to facilitate easy removal for work on smaller specimens.
Component Masses and Positions. Volumes were obtained from the model for the shell, siphuncle and body chamber using functions built into the software (Table 3).
 The total volume of the septa was calculated as the product of the septal face surface areas times thicknesses. As discussed for the septum in model 1, the volumes of the septa and shell were reduced by 7.6 % before being included in the model to account for the change of volume caused by aragonite inverting to calcite (Table 3). To obtain the volume of cameral sheets, total chamber volume was calculated by subtracting the volume of the septa from the total volume of the interior of the phragmocone obtained from the model. As in model 1, the volume of cameral sheets was included at 4.5 % offset with a total of 9.5 % of chamber volume being filled. Similar to model 1, the density used for the septa and shell was 2.67 g/cm3 (Hewitt and Westermann 1996) whereas the density used for the siphuncle, cameral sheets and body chamber was 1.055 g/cm3 (Saunders and Shapiro 1986). Density multiplied by volume provided the mass of each component (Table 3). The position of the centroid of each individual component in the X, Y and Z plane with respect to the origin is given in
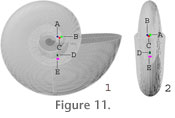
Table 3 (Figure 11). The origin (X, Y, Z = 0, 0, 0) is the very centre of the model. In the innermost whorl, the position of the siphuncle is variable but by the next whorl the position of the siphuncle is reasonably stable, and the suture line is consistently expanded on the non-siphuncle side and reduced on the siphuncle side. The total volume of the septa was calculated as the product of the septal face surface areas times thicknesses. As discussed for the septum in model 1, the volumes of the septa and shell were reduced by 7.6 % before being included in the model to account for the change of volume caused by aragonite inverting to calcite (Table 3). To obtain the volume of cameral sheets, total chamber volume was calculated by subtracting the volume of the septa from the total volume of the interior of the phragmocone obtained from the model. As in model 1, the volume of cameral sheets was included at 4.5 % offset with a total of 9.5 % of chamber volume being filled. Similar to model 1, the density used for the septa and shell was 2.67 g/cm3 (Hewitt and Westermann 1996) whereas the density used for the siphuncle, cameral sheets and body chamber was 1.055 g/cm3 (Saunders and Shapiro 1986). Density multiplied by volume provided the mass of each component (Table 3). The position of the centroid of each individual component in the X, Y and Z plane with respect to the origin is given in
Table 3 (Figure 11). The origin (X, Y, Z = 0, 0, 0) is the very centre of the model. In the innermost whorl, the position of the siphuncle is variable but by the next whorl the position of the siphuncle is reasonably stable, and the suture line is consistently expanded on the non-siphuncle side and reduced on the siphuncle side.
 Thus, we considered the offset distances of the siphuncle, septa and cameral sheets to be the same as in model 1 (Table 2). Finally, the data for each component was combined, and centres of buoyancy and mass were calculated for the entire ammonite animal. The centres of mass (total X, Y and Z coordinates in
Table 4) and buoyancy (displacement in
Table 4) are shown in
Figure 12. The final X, Y and Z coordinates are calculated from the total weighted coordinate for each axis divided by the total mass of the animal. Thus, we considered the offset distances of the siphuncle, septa and cameral sheets to be the same as in model 1 (Table 2). Finally, the data for each component was combined, and centres of buoyancy and mass were calculated for the entire ammonite animal. The centres of mass (total X, Y and Z coordinates in
Table 4) and buoyancy (displacement in
Table 4) are shown in
Figure 12. The final X, Y and Z coordinates are calculated from the total weighted coordinate for each axis divided by the total mass of the animal.
Model 3: Entire Ammonite Animal at 54 mm Maximum Shell Diameter
To investigate how these asymmetries affected the entire ammonite animal at different growth stages, a three-dimensional model of a complete, small ammonite animal was also created (model 3, DMAX = 54 mm). Model 3 was constructed by removing the outer two whorls from model 2 and altering the position of the body chamber. The volumes were calculated using the same methods described above for model 2, and the densities used for each component are the same as in model 2 (Table 5). Volumes, masses and relative positions of the centroids of each component in the X, Y and Z plane are given in
Table 5 whereas the positions of the centres of buoyancy (displacement) and mass (final X, Y and Z coordinates) are provided in
Table 6. Once again, cameral sheets are kept optional.
|