|
|
|
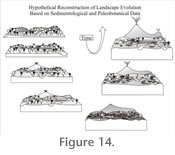
DiscussionDespite the frequent and widespread occurrence of Cenozoic volcaniclastic deposits, fine-scale paleoenvironmental reconstruction through a continuous geological section has been attempted only in a few instances (e.g., Yuretich 1984; Crowley et al. 1989, 1994; Falcon-Lang and Cantrill 2002; Jago and Boyd 2005). Most studies have focused on single sedimentary events that represent a snapshot of the plant community at a given moment. In these cases, the dynamics of plant community changes as influenced by concomitant changes in the environment, particularly by volcanic activity, remain unsampled. Moreover, inferences of environmental conditions based on data from a single sedimentary event is assumed to be a good representative of landscape heterogeneity at the time of deposition, but may be misleading in rapidly changing regions, such as those influenced by catastrophic events. The study presented here is among the few that have used a combined approach, including sedimentology, stratigraphy, and paleobotany, to infer the evolution of the environment at fine temporal and spatial scales, and its focus on the poorly known Paleogene of tropical Africa makes it unique. Interpretation of SedimentologyThe main characteristic of the strata represented in the sections studied here is that they are thin, incompletely/poorly-developed, and often randomly truncated or mantled by massive ash layers. These attributes most likely resulted from stochastic variation in sediment discharge influx and therefore irregular rates of deposition. In particular, sedimentary structures present in these strata can be related to fluvial processes associated with channel and overbank depositional environments (e.g., Currie et al. 2009). The distinction between these depositional environments is not sharp, but gradual since the sedimentary structures concerned represent adjacent areas of the same fluvial environment. Characteristics that help to distinguish between these depositional environments are the association of particular sedimentary structures and the grain size of the sediments (e.g., Currie et al. 2009). In this study, most depositional units are formed of fine-grained particles, which indicate that they represent overbank and nearby areas of a fluvial environment. The lower portion of the stratigraphy, named stage 1 in Figure 3, consists of fine-grained sediment and organic-rich layers (E, F depositional units; all beneath Ash-IV), classified here as a Vertisol paleosol, suggesting a relatively long depositional hiatus in the overbank depositional environment. Emplacement of the stage 1 strata are laterally continuous among the geological sections, and the abundant organic remains (e.g., leaves) concentrated in a thin horizon capping the sequence probably represent forest litter accumulation. The overlying stage 2 strata (Figure 3; beginning of Ash-IV deposition) consist of weakly-laminated claystones, mudstones, and siltstones (D, E, F depositional units), and include fossil leaves and root traces that are most abundant in a thin horizon at the base of the sequence. Stage 2 sediments are interpreted as a Protosol paleosol created by suspension-settling after flooding in a vegetated overbank environment relatively distant from the effects of fluvial-channel depositional processes. Both stage 1 and 2 strata (Figure 3) are consistent with fluvial deposition characterized by meandering streams (Thomas et al. 1987), whereas the latter probably represents a shorter time interval and closer proximity to the channel environment based on generally coarser-grained and relatively lower organic content of the sediments. The thin organic-rich horizon at the base of the stage 2 strata also probably represents forest litter accumulation, but for a shorter period of time relative to stage 1. Stage 3 (Figure 3) includes organic-rich, poorly developed cross- and horizontally-laminated fining upward strata that represent partially preserved heterolithic cross stratification (A, C, D, and E depositional units). Stage 3 is interpreted to represent parts of point-bar deposits associated with lateral accretion processes in a fluvial environment (Thomas et al. 1987). However, bedding contact surfaces within this succession can be hummocky to erosional, suggesting that these strata may alternatively represent a succession of thin levee deposits in overbank environments characterized by deposition from a meandering stream. Stage 4 (Figure 3) consists of relatively thicker, massive to poorly-laminated and ripple-cross-laminated, siltstones and very fine sandstones (C, B depositional units) formed by gravity-flow deposition. This deposition mode commonly occurs in channels when overbank slumping results in the sudden formation of a sedimentary deposit (Miall 1996; Boggs 2006). However, the strata considered here consist mainly of volcanic ash that was not already part of the system, but rather was supplied by local to regional volcanic eruptions. This indicates that part of the extra sediment supply released into the environment was directly deposited on parts of the floodplain, while some of it choked streams and promoted sluggish sedimentation. As a result, sedimentary features characterizing these tuffaceous strata are associated with the channel/overbank transitional part of the depositional environment, much like the preceding units. Stage 5 consists of poorly-developed (i.e., partially unconsolidated organic matter) lignites and other poorly-laminated organic-rich layers (e.g., Figure 3, CH-93, unit 11-13). The majority of these strata are classified within the fine-grained and organic-rich depositional units (D, E, F, and G) and therefore they represent sedimentary structures typical of the overbank depositional environment (Miall 1996). Their presence is consistent with autochthonous and parautochthonous deposition of organic matter in ephemeral ponds or other parts of the floodplain characterized by a high water table (Miall 1996; Boggs 2006). The origin of these ephemeral pools of standing water can be the result of the obliteration of topographic differences and sealing of soils and other surfaces on the floodplain by the episodic deposition of volcanic ash (Crowley et al. 1989, 1994). Stage 6 includes cross-laminated, thin, fine sandstones which scour underlying sediments of succession 5 and are, in turn, overlain by massive to poorly-laminated finer-grained layers. This upper part of the succession also shows increasing bed thickness and decreasing organic content toward the north section (CH-89). A thin and continuous organic-rich clayey layer caps the succession. The more coarsely laminated layers are interpreted as fluvial deposits associated with flash flood events, while the massive to poorly reworked strata probably represent rapidly deposited airfall to stream-deposited tuffs buried in a more or less proximal position on a floodplain. The laminated strata represent sedimentary structures formed in the overbank environment; in particular, based upon their gradual thickening and development of scour sedimentary structures in a south (CH-79) to north direction (CH-89), these sedimentary structures represent transition from lower levee deposits to crevasse channel deposits in a position proximal to stream channel environments (Miall 1996). Additional massive to poorly-laminated siltstones, some of which also show a concave shape, akin to scour and fill structures, characterize stage 7. These have been assigned to the C, B, and D depositional units and are interpreted as sedimentary structures formed or deposited in the overbank environment (Miall 1996). In particular, the horizontal to wavy laminated layers and the layers showing scour and fill structures represent various transitional parts ranging from the overbank to the channel depositional environments, respectively (Miall 1996). The massive siltstones are interpreted to have been aerially deposited on different parts of the fluvial landscape, where they created deposits on the floodplain and choked streams resulting in the formation of multiple small channels according to a braided pattern. In summary, from the sedimentological data analyzed, it is inferred that most subenvironments of a fluvial environment, except for the most distal parts of the floodplain, are preserved within the succession of strata associated with the Ash-IV unit and the thin set of strata below. An overview of the sedimentary structures of the deposits analyzed is consistent with initial (pre-AshIV strata) deposition by meandering streams on a seasonally inundated floodplain. With the beginning of Ash-IV deposition, calm sedimentation was interrupted by rapid accumulation of ash on different parts of the landscape. Numerous thin lignitic horizons intercalated with these sediments in the stratigraphic middle part of the sections indicate in situ deposition of organic matter in ephemeral pools of stagnant water distributed on the floodplain during volcanic quiescence. Increased frequency of airfall and fluvially reworked ashes, in the upper part of all stratigraphic sections, indicate environments characterized by ash-covered floodplains and fluvial deposition according to a braided pattern. The distribution of depositional units and paleosols indicating the rapid evolution of a system characterized by rather calm deposition on seasonally inundated floodplains to more energetic deposition by braided streams is consistent with stochastic variation of sedimentary discharge rates, which probably reflects the increasing influence of episodic volcanic activity on the landscape through time. Interpretation of Plant AssemblagesAmong the fossil plants preserved, angiosperms occur throughout the basalmost layers of Ash-IV and the thin set of strata below (pre-Ash-IV) with leaf remains concentrated in two thin horizons probably representing forest litter accumulation. Numerous compressions and impressions of, apparently, several different angiosperm morphotypes occur in these basal horizons, but most of these are impossible to differentiate, since they overlap and are tightly appressed, thus precluding the possibility of identification. The most abundant fossil in these two horizons and the only taxon present in the overlying layer of Ash-IV in all sections is Cynometra, which is also present in stratigraphically older strata of the Chilga beds (Pan 2007). There, Cynometra is part of a paraautochthonous plant assemblage that was interpreted as representative of a riparian forest environment (Pan 2007) and it is reasonable to assume the same is true in this case.
Plant communities adjacent to or within depositional areas above the basal horizons characterized by forest litter accumulation appear less rich taxonomically up section, where angiosperms are generally underrepresented and ferns dominate. Fossil ferns have been shown to be the principal component of low diversity plant communities in other studies of environments subject to disturbance by volcanic activity (Jago and Boyd 2005;
Lentfer and Torrence 2006). In these
communities, dominance by ferns has been attributed to their role as ecological pioneers (Went 1959;
Richards 1996). A number of studies have shown that volcanic ash deposition in fluvial environments can cause sealing of soils and choking of rivers, promoting flooding and consequent erosion and runoff (Wilcox 1959; Burnham and Spicer 1986). Other authors have indicated that leaching and deposition of volcanic ashes also results in impermeable horizons, which promotes pond development (Crowley et al. 1989, 1994). Pond development is associated with a change of vegetation, where ferns can dominate in the early stages (Cohen and Raymond 1984). The abundance of ferns may also be attributed to their capacity to live in poorly oxygenated, waterlogged settings, as well as their ability to be early ecological colonizers (Wing et al. 1993; Page 2002). Accordingly, the abundance of ferns in all the lignites and similar organic rich layers is interpreted as an occurrence of ephemeral ponds across the sampled area in response to volcanic influence on the landscape and the ability of this plant group to live in this kind of setting. The possibility that the occurrence of more fern remains in the lignitic/peaty layers relative to angiosperms was the result of taphonomic processes should also be considered. Spicer (1989) suggested that reed vegetation growing around the margins of swamps could form a natural barrier preventing entrance and sedimentation of water-transported plant fragments from elsewhere. The few angiosperm remains preserved in the Ash-IV lignites could have been windblown. However, there is a consistency in the composition of the ferns and angiosperms, which are always disturbance taxa (e.g., Macaranga and Cyclosorus) (Slik and van Welzen 2001). These occur immediately above the lignitic/peaty horizons in some of the massive to slightly reworked tuffs, which are interpreted as open ash-covered landscapes created by volcanic activity. As a result, it seems more plausible that ferns and angiosperms characterizing both lignites and ash layers are a reflection of local low diversity plant communities rather than a consequence of taphonomic processes restricting the preservation or input of angiosperms. This interpretation is consistent with previous research showing that plant communities subjected to recurrent volcanism are characterized by poor taxonomic diversity (Jago and Boyd 2005; Lentfer and Torrence 2006). Interestingly, up-section, the ferns from Chilga often co-occur with several palms (including Calamoid and Coryphoid taxa), a plant association documented before in paleoenvironments directly influenced by volcanic activity (Wing et al. 1993). Some palms are known as early colonizers in successions from herbaceous to forest swamps, and where they are adapted to saline conditions (Coetzee and Rogers 1982; see Collinson 2001 for other examples where an early successional role has been attributed to palms). Although sediments from the Margargaria River localities do not indicate saline conditions, the association of palms with some or all of the fern taxa (Acrostichum, Blechnum, Cyclosorus, Equisetum, Marsilea, Salvinia) interpreted here as early ecological colonizers is consistent with a similar role for palms. Previous research on palms from Chilga documents their occurrence in a variety of sediment types representing similar and different edaphic settings compared to those described here, which is consistent with ecological versatility and the ability of palms to live in rapidly changing environments (Pan et al. 2006). Plant assemblages described in this study appear compositionally and ecologically unique when compared to other Paleogene communities from Africa (see e.g., Jacobs 2006; García Massini and Jacobs 2009; Jacobs et al. 2009). This appears evident considering the scale of interpretation of the Margargaria localities analyzed here, the scarcity and uneven distribution of Paleogene macrofloras in Africa, and the poor age control of most known plant assemblages for this period (see e.g., Jacobs 2006; García Massini and Jacobs 2009; Jacobs et al. 2009). Nevertheless, some of the plant taxa present in the Margargaria River localities are also found in similar environments from the Paleogene in Africa. For example, Acrostichum, Salvinia, and Typha macrofossils are known from possible Eocene coastal swampy/marshy environments from Nigeria (Seward 1924). These same three taxa are known from the Late Eocene of Egypt (Fayum depression) in what are interpreted as mangrove and brackish (Acrostichum) and marshy habitats (Salvinia and Typha), while other taxa in the same deposits, characterized by angiosperms that are not found in the plant assemblages from the Margargaria River, indicate gallery forests inland (Tiffney 1991). Acrostichum macrofossils are also known from Uganda (Middle Tertiary) and Ethiopia (Oligocene; to the south of Chilga) in woodlands or savannahs and in lacustrine deposits, respectively, both set in volcanic environments (Chaney 1933; Beauchamp et al. 1973). Co-occurring with Acrostichum in the two latter places are a number of dicot angiosperm taxa, none of which, except for the presence of the family Fabaceae, are present in the Margargaria River localities (Chaney 1933; Beauchamp et al. 1973). In fact, the Fabaceae appears to be the most abundant family preserved as macrofossils (leaves and wood) and palynomorphs in African Cenozoic assemblages, although the genus Cynometra, which characterizes some of the strata from the Margargaria River, is only known from younger strata of Chilga and from Middle Eocene deposits from Tanzania (Jacobs 2006; Pan 2007). In this latter deposit, Cynometra occurs as part of a legume-dominated plant assemblage characterized as a seasonal woodland environment (Herendeen and Jacobs 2000). This is to some extent consistent (i.e., both represent seasonal forest environments) with the depositional environment interpreted for the occurrence of Cynometra from the Margargaria localities. Other than these four taxa, the Margargaria River flora shares a record of Pandanus (seeds) from the Fayum depression (Kräusel and Stromer 1924), but this evidence has been questioned by Chandler (1954). Palynologically, Macaranga-type pollen and Acrostichum, Salvinia, and Marsilea spores have been recorded from a series of settings ranging from coastal mangrove and swampy settings to inland semi-deciduous forest with humid woodland West and East Africa (e.g., Caratini et al. 1991; Vincennes et al. 2006); however, identification of palynomorphs relative to macrofossils, especially for dispersed spores, is less precise due to fewer diagnostic characters (e.g., see Collinson 2001, 2002). Moreover, edaphic settings for those assemblages are not comparable to the Margargaria River, where volcanism was the driver for the occurrence and rapid deposition of plant assemblages. As a result, the scarce representation of the macrofossils from the Margargaria River at other Paleogene localities in Africa and the particular setting that volcanism associated with the development of the East African Rift represents make the Ethiopian plant assemblages considered in this study unique. Notwithstanding, some of the taxa present in the Margargaria River localities, such as Acrostichum, Salvinia, and Typha, are also found to be common in similar settings (swamp/marshy habitats, sometimes influenced by volcanism) in other African Paleogene deposits, which may be indicating ecological consistency for these taxa during this period in this continent. These same taxa, as well as Blechnum, Cyclosorus, and various palms are also common, single or in combination, throughout North America-Europe during the Paleogene. This supports the common association of these taxa in similar habitats characterized as wetland that were in some instances directly influenced by volcanism during the Paleogene (Collinson 2001, 2002 and references therein). In summary, the Ash-IV deposits and the thin set of strata below pre-Ash-IV) contain a succession of compositionally and ecologically unique plant communities that are a reflection of the physiographic characteristics of the environment. In particular, the angiosperm taxa characterizing the early depositional stages of Ash-IV, as well as the thin set of strata below (pre-Ash-IV), correspond to riparian forest vegetation typical of seasonal environments. Taxa in the middle and upper part of the geological sections can be characterized as ecological pioneers, where their presence reflects the influence of rapid and episodic modifications of the landscape triggered by volcanic activity. Volcanism provided an inorganic, fresh surface of ash for colonization by pioneer plants and a sealant that preserved the pre-existing vegetation at the time of each eruption. This indicates that, as the physical settings within the environment were episodically disturbed, so were plant communities reset to earlier stages of development and, in this sense, these deposits preserve several stages of plant recovery, particularly those stages associated with early colonization. This is consistent with low diversity plant assemblages and the mode of preservation and size disparity of plant parts preserved, especially in the airfall tuffs, all of which indicate poor sorting and rapid deposition. Therefore, plant taxa in the middle and upper parts of the studied sections can be interpreted as the fittest available vegetation source following each particular volcanic eruption rather than the temporally-directed (undisturbed by major catastrophic events) evolution of a single plant community through ecological succession. Summary of Paleoenvironmental Stages Based on Paleobotany and SedimentologyStratigraphic correlations among the three sections described from Ash-IV and the thin set of strata below delineate a general evolutionary pattern of ecosystem change through time (Figure 3, Figure 14.1-14.7). Rather calm sedimentary conditions, dominated by a meandering river, correspond to the initial depositional stages (pre-Ash-IV) (Figure 3, Figure 14.1). During the second to third stages (beginning of Ash-IV deposition) (Figure 3, Figure 14.2-14.4) increased deposition rates result from intermittent ashfall. Some of this ash was aerially deposited, as is indicated by the presence of massive strata, and some was reworked and deposited fluvially. The upwards increasing presence of fill and scour structures internally cross-laminated to lenticular-bedded implies continuous ingression of ash that rapidly filled and choked streams with sediment. These structures indicate that small-scale braided streams, instead of large meandering streams gradually developed on the landscape. Apparent sedimentary calm is inferred from the presence of vertically and laterally alternating lignites and organic-rich strata dominated by ferns (Figure 3, Figure 14.5). This indicates widespread low-lying areas where deposition of organic material occurred in situ. The presence of ashy material within these deposits indicates that ashes covered a vast part of the landscape, and that intermittent fluvial deposition reworked some of this ash into environments characterized by peat accumulation. With time, volcanism gradually resumed and fluvial sedimentation occurred by means of numerous small penecontemporaneous channels in a braided stream system. This relatively unstable environment resulted in fewer plants able to occupy the environment and in situ organic accumulation became less common across this region of the landscape (Figure 3, Figure 14.6). Increased abundance of massive tuffs up section indicates active volcanic activity, while the vertical repetition of scour and fill structures, intercalated with the ashy horizons, is evidence of the sporadic deposition by means of high energy braided streams (Figure 3, Figure 14.7). The abundance of ferns and a few other pioneer colonizers in some of these strata is consistent with an unstable environment, where only those plants adapted to the rapidly changing physiography were able to endure. In summary, the data presented here show that episodic volcanic activity directly influenced the evolution of the environment and plant community, where a forested landscape characterized by rather calm fluvial deposition was replaced by one dominated by ferns and characterized by aerial and dynamic fluvial deposition. |
|