FIGURE 1. Basal protomonaxonids and a primitive hamptoniid from the early Cambrian Hetang Biota, South China. 1-2, NIGP154828 Hyalosinica sp.; 1, overall view with examples of hexactines arrowed; 2, magnification of hexactines in central part of (1); 3, NIGP155891 apparently undescribed genus (new genus B) with mass of minute, inflated and short-rayed hexactines (visible as tiny shadowed cavities) amongst irregularly-arranged, fine monaxons; 4-5, NIGP155892 undescribed genus (new genus A) with dense wall of minute, inflated-rayed hexactines and no monaxons; 4, overall view showing body form; 5, detail showing mass of minute, short-rayed hexactines preserved as external moulds; 6, NIGP155893 probably undescribed genus (new genus C) with irregular, dense array of sub-longitudinal monaxons but lacking hexactine-based spicules, interpreted as a primitive hamptoniid-like sponge due to the lack of hexactine-based spicules; 7-8, NIGP155894 probably undescribed genus with conical body wall containing mostly diagonally-oriented monaxons, and non-inflated hexactine-based spicules. Scale bars equal: 1, 3: 5 mm; 2, 5, 8: 1 mm; 4, 6-7: 10 mm.

FIGURE 2. Leptomitidae, from the Burgess Shale (middle Cambrian, Canada).1, 3 Leptomitus lineatus ROM 53558; 1, apex of sponge showing longitudinal monaxon bundles converging towards narrowed osculum, and short transverse monaxons; 3, basal region with linear skeletal nucleation centre; 2, 4 Wapkia elongata; 2, ROM53549, showing fundamental architecture of longitudinal spicules with prominent development of transverse monaxon bundles, distorted into plumose array in left half; 4, ROM 53544, apical region of holotype showing skeletal architecture. 5, 7, ROM6l9l5, probably undescribed leptomitid-like sponge with thick wall and complex architecture from Burgess Shale locality S7; 6, ROM6l9l0, undescribed protomonaxonid from the Burgess Shale locality S7; previously named as Leptomitus but lacks transverse spicules (photograph: J.-B. Caron, ROM). 1-3, 5, 7 photographed under crossed polarisers, with high-angle illumination; 4 photographed with low-angle illumination. Scale bars equal: 1, 7: 1 mm; 2-5: 5 mm; 6: 50 mm. ].

FIGURE 3. Hamptoniidae and 'Choiidae'. 1, Hamptonia bowerbanki, ROM6l9l4 from Monarch Cirque, British Columbia (ROM), with bimodal array of monaxons including relatively large, prominent spicules embedded in finer thatch; 2-3, NIGP155895 Choia sp. from the Hetang Biota; 2, overall view showing projecting monaxons; 3, magnified view of central body showing acicular coronal spicule bases (compare with Figure 4); 4, Lenica sp. NIGP154161, detail showing conical, organic-walled spicules preserved as flattened, partly pyritised moulds. Scale bars equal: 1: 10 mm; 2, 4: 5 mm; 3: 2 mm.

FIGURE 4. Halichondritid and piraniid sponges from early and middle Cambrian Lagerstätten. 1,3, apparently undescribed halichondritid-like sponge NIGP155896, Hetang Biota (early Cambrian, Anhui, South China); 1, view of partial specimen, showing architecture of fine monaxons and open-based, conical spicules; 3, detail with single conical spicule; 2, 4, Choia hindei (ROM53563) from the Burgess Shale, showing bimodal spicule sizes, the flattened open bases of large spicules shown in D; 5-6 Pirania muricata; 5, ROM6l9l3, details of central parts of spicules with preserved axial regions, dissolved middle layer, and organic preservation of outer film; 6, ROM53589, detail showing insertion of spicules into sponge wall, with poorly-defined, apparently open bases. Scale bars equal: 1-2: 5 mm; 3, 5-6: 1 mm; 4: 2 mm.

FIGURE 5. Hazeliidae from the Burgess Shale (middle Cambrian, British Columbia, Canada). 1, Hamptoniella foliata ROM44283, with sub-longitudinal array of spicule tracts and irregular cross-branching and reticulation; 2, Hazelia palmata ROM56247, distal part of large frondose specimen showing continuation of spicule tracts beyond margin; 3, ROM6l9l2, an undetermined hazeliid with a moderately thick, but disorganised wall and no reticulation; 4-5, ROM53578, new taxon described as Crumillospongia frondosa (Rigby and Collins, 2004) but showing echinating spicule tracts incompatible with that genus, visible in both cross-polarised (4) and low-angle light (5); 6-7, Hazelia delicatula ROM56258, with detail of upper right region (7) showing reticulate skeleton. Scale bars equal: 1, 6: 5 mm; 2, 4-5, 7: 1 mm; 3: 10 mm.

FIGURE 6. Members of the vauxiid-anthaspidellid complex, from the Burgess Shale (middle Cambrian, British Columbia, Canada). 1, detail of Hazelia conferta ROM56253, with sub-regular reticulation of primary skeleton; 2,4, Vauxia cf. bellula ROM6l 9l l, showing regular hexagonal (pseudo-rectangular) reticulation of longitudinal columns, lateral parts complicated by superposition of the dermal layer, and detail (4) showing framework composed of discrete, usually paired and sometimes misaligned spicules; 3, Vauxia bellula ROM 56243; 5, Fieldospongia bellilineata ROM53602, showing regular arrangement of single and double monaxons to form hexagonal (sub-rectangular) reticulation; 6, undescribed fragment of anthaspidellid lithistid on same slab as ROM44283, with more complex skeletal architecture. Scale bar equals 1 mm.

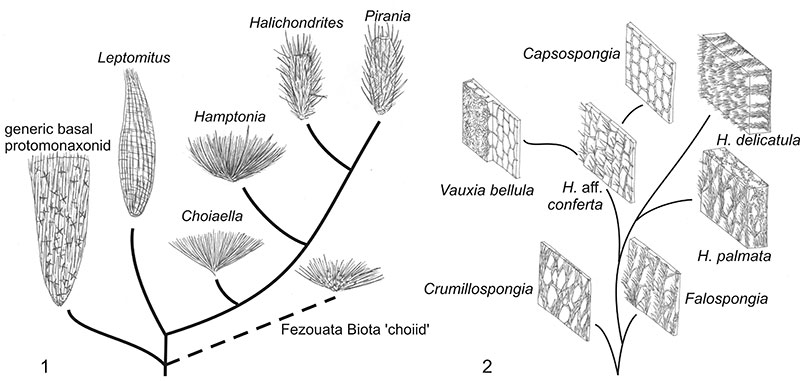
FIGURE 7. Phylogenetic hypothesis for the protomonaxonids, divided into two major groups: 1, large-spiculed taxa, including links to basal, hexactine-bearing sponges; 2, small-spiculed taxa with relationships to modern demosponge lineages. New genera A, B, and C are described briefly in the text. This reconstruction includes most of the genera described from Cambrian deposits, but a few poorly-understood or potentially problematic taxa are excluded. Vertical axis represents evolution; no scale of time or morphology is implied.
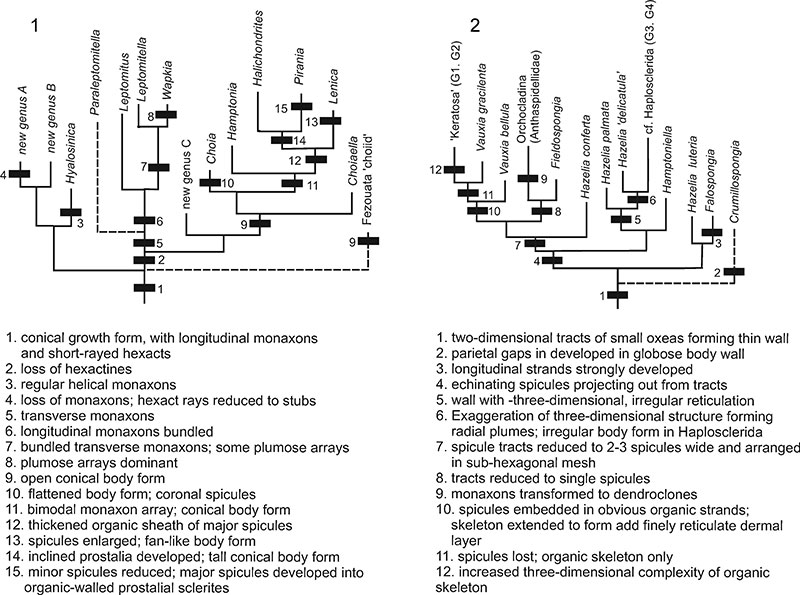
FIGURE 8. Schematic illustration showing primary skeletal development in protomonaxonids, referring to the most fundamental changes described in Figure 7.1, Group One (refers to Figure 7.1); 2, Group Two (refers to Figure 7.2).