

APPENDIX 1.
Terms, definition, and usage of pneutmatic morphology and histology used in this analysis.
Pneumatic Architecture (Wedel et al., 2000)
Camera - Round cavity, 5-150 mm in size, septal thickness of 2-10 mm, and a regular branching pattern.
Camella - Angular cavity, 2-20 mm in size, septal thickness of 1-3 mm, and an irregular branching pattern.
Acamerate - Pneumatic structures limited to fossae. Fossae do not significantly invade the centrum.
Procamerate - Deep fossae penetrate to median septum, but are not enclosed by osteal margins.
Camerate - Large, enclosed camerae with regular branching pattern; cameral generations usually limited to 3.
Camellate - Fine internal structures composed entirely of small scaled, thin-walled camellae; can produce a “honeycomb”-like network.
Histologic Terms (Francillon‐Vieillot et al., 1990; de Ricqlès et al., 1991; Castanet et al., 1992; Huttenlocker et al., 2013)
Line Of Arrested Growth (LAG) - Thin bands that represent temporary arrest of osteogenesis, and are considered osteological response to predictable environmental cues.
Annuli - Translucent to opaque bands, thicker than LAGs, represent a slowing (but not a cessation) of osteogenesis.
Woven Bone - Highly disorganized arrangement of collagen fibers, which reflects a high rate of osteogenesis.
Fibrolamellar Bone - Woven bone with intervening and randomly oriented primary osteons.
External Fundamental System (EFS) - Slowly deposited parallel-fibered or lamellar tissue along the outermost cortex (closely spaced outermost series of LAGs).
Haversian Bone - Bone that is completely remodeled by secondary osteons.
Primary Osteon - Central blood vessel and surrounding concentric bone tissue.
Secondary Osteon - Osteon formed by replacement of existing bone, surrounded by an outer cement sheath.
Cancellous Bone - Highly vascular bone that contains a higher surface area to mass ratio.
Trabeculae - Rod-shaped bone tissue in cancellous bone. Provides lightweight internal support.
Laminar Vascular Canal - Circumferentially oriented rows of vascular canals.
Longitudinal Vascular Canal - Canals oriented parallel to the long axis on the bone.
Reticular Vascular Canal - Obliquely oriented vascular canals.
APPENDIX 2.
Sauropod specimens examined in this analysis (H-MOS = Histo-Morph Ontogeny Scale [see Appendix 3]).
| Specimen Number | Material Examined | Taxonomy | H-MOS Estimation |
| AMNH 353 | Femur | Apatosaurus sp. (McIntosh, 1990) | Stage 4 |
| AMNH 435 | Femur | Apatosaurus sp. (this analysis) | Stage 2 or 3 |
| AMNH 606 | Femur | Apatosaurus sp. (this analysis) | Stage 4 |
| AMNH 613 | Femur | Diplodocidae (this analysis) | Stage 1 |
| AMNH 5855 | Femur | Diplodocus sp., Barosaurus (Mook, 1917; McIntosh, 2005) | Stage 2 |
| AMNH 6341 | Vertebrae | Barosaurus (Lull, 1919; Tschopp et al., 2015) | Stage 3 or 4 |
| AMNH 7530 | Vertebrae | Barosaurus, Kaatedocus (Michelis, 2004; Tschopp et al., 2015) | Stage 2 |
| AMNH 7535 | Vertebrae | Barosaurus (Michelis, 2004; Tschopp et al., 2015) | Stage 2 or 3 |
| AMNH 7539 | Femur | Diplodocus sp. (this analysis) | Stage 2 |
| ANS 21122 | Vertebrae, Femur, Skull, Dorsal Ribs | Suuwassea, Apatosaurinae, Dicraeosauridae (Harris, 2006a,b; Whitlock and Harris, 2010; Woodruff and Fowler, 2012; Tschopp et al., 2015) | Stage 3 |
| BYU 601-17103 | Femur | Apatosaurus sp. (Wilhite, 2003) | Stage 4 |
| BYU 725-4889 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 2 |
| BYU 725-9026 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 2 or 3 |
| BYU 725-11421 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 2 or 3 |
| BYU 725-12155 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 3 |
| BYU 725-13369 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 3 |
| BYU 725-13643 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 4 |
| BYU 725-13670 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 1 |
| BYU 725-16569 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 2 |
| BYU 725-16610 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 2 |
| BYU 17096 | Skull | Apatosaurus sp. (Balanoff et al., 2010) | Stage 2 |
| BYU Fe-4-DM197 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 3 |
| BYU Fe-5-DM172 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 3 |
| CM 84 | Vertebrae, Femur | Diplodocus carnegii (Hatcher, 1901; Tschopp et al., 2015) | Stage 4 |
| CM 85 | Femur | Apatosaurus sp. (McIntosh, 1981) | Stage 4 |
| CM 87 | Femur | Apatosaurus sp. (McIntosh, 1981) | Stage 4 |
| CM 94 | Vertebrae, Femur, Dorsal Ribs | Diplodocus carnegii (Hatcher, 1901; Tschopp et al., 2015) | Stage 4 |
| CM 555 | Vertebrae | Apatosaurus excelsus,Brontosaurus excelsus (McIntosh, 1981; Tschopp et al., 2015) | Stage 3 |
| CM 563 | Vertebrae | Apatosaurus excelsus, Brontosaurus parvus (Gilmore, 1936; Tschopp et al., 2015) | Stage 4 |
| CM 566 | Femur | Apatosaurus sp., Brontosaurus parvus (McIntosh, 1981; Tschopp et al., 2015) | Stage 1 |
| CM 572 | Vertebrae | Haplocanthosaurus priscus (Hatcher, 1903; Tschopp et al., 2015) | Stage 3 or 4 |
| CM 879 | Vertebrae | Haplocanthosaurus utterbacki (Hatcher, 1903; Tschopp et al., 2015) | Stage 2 or 3 |
| CM 3018 | Vertebrae, Femur, Skull | Apatosaurus louisae (Gilmore, 1936; Tschopp et al., 2015) | Stage 4 |
| CM 3390 | Vertebrae | Apatosaurus sp. (McIntosh, 1981) | Stage 2 |
| CM 11161 | Skull | Diplodocus longus; Diplodocinae Indeterminate (Berman and McIntosh, 1978; Tschopp et al. 2015) | Stage 4 |
| CM 11162 | Skull | Apatosaurus louisae (Berman and McIntosh, 1978; Tschopp et al., 2015) | Stage 4 |
| CM 11338 | Vertebrae | Camarasaurus lentus (Gilmore, 1925) | Stage 2 |
| CM 21785 | Femur | Apatosaurus sp. (McIntosh, 1981) | Stage 4 |
| CM 21788 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 1 or 2 |
| CM 30762 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 2 |
| CM 30766 | Femur | Apatosaurus sp., Apatosaurine (Wilhite, 2003; Tschopp et al., 2015) | Stage 3 |
| CM 33976 | Femur | Apatosaurus sp., Diplodocus sp. (Wilhite, 2003, this analysis) | Stage 1 |
| CM 33991 | Femur | Diplodocus sp. (McIntosh, 1981) | Stage 1 |
| CMC VP 7747 | Femur | Diplodocidae, Diplodocus sp. (Meyers, 2004; Woodruff and Fowler, 2004) | Stage 2 |
| CMC VP14128 | Skull | Diplodocus sp. (this analysis) | Stage 2 |
| CMNH 10039 | Femur | Apatosaurus sp. (Wilhite, 2003) | Stage 2 |
| CMNH 10380 | Vertebrae | Haplocanthosaurus delfsi (Wilhite, 2003) | Stage 4 |
| DNM 3781 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 4 |
| GPDM 220 | Vertebrae | Camarasaurus sp. (this analysis) | Stage 4 |
| HMNS 175 | Femur | Diplodocus hayi, Galeamopus (Holland, 1924; Tschopp et al., 2015) | Stage 4 |
| KUVP 1351 | Femur | Apatosaurus sp. (Wilhite, 2003) | Stage 4 |
| MOR 592 | Vertebrae, Femur, Skull, Dorsal Ribs, Cervical Ribs | Amphicoelias altus, Dicraeosauridae, Diplodocus sp. (Wilson and Smith, 1996; Whitlock and Harris, 2010; Woodruff and Fowler, 2012) | Stage 3 |
| MOR 700
(2 specimens) |
Skull, Cervical Ribs | Apatosaurus sp. (Woodruff and Fowler, 2014) | Stage 2 and 4 |
| MOR 714 | Vertebrae | diplodocid indeterminate (this analysis) | Stage 1 |
| MOR 790
(15 specimens) |
Vertebrae, Femur, Dorsal Ribs, Cervical Ribs | Diplodocinae; Diplodocus sp. (Myers, 2004; Woodruff and Fowler, 2012) | Stage 2 |
| MOR 957 | Vertebrae, Femur | Apatosaurus sp. (this analysis) | Stage 4 |
| MOR 7029 | Vertebrae, Skull | Diplodocus sp. (Woodruff and Fowler, 2014) | Stage 3 |
| MWC 5439 | Femur | Apatosaurus sp. (this analysis) | Stage 1 |
| MWC
“Moffit Co. Apato.” |
Femur | Apatosaurus sp. (this analysis) | Stage 4 |
| NSMT-PV 20375 | Vertebrae, Femur | Apatosaurus ajax, Apatosaurinae Indeterminate (Upchurch et al., 2004b; Tschopp et al., 2015) | Stage 4 |
| OMNH 1793 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 1 |
| OMNH 01667 | Femur | Apatosaurus sp. (Wilhite, 2003) | Stage 4 |
| SMA 0003 | Vertebrae | Diplodocidae, Diplodocus sp. (Schwarz et al., 2007a; this analysis) | Stage 3 |
| SMA 0004 | Vertebrae, Skull | Kaatedocus (Tschopp and Mateus, 2012; Tschopp et al., 2015) | Stage 2 or 3 |
| SMA 0009 | Vertebrae, Femur, Cervical Ribs | Diplodocidae; Barosaurus, Brachiosaurus (Schwarz et al., 2007a; Woodruff and Fowler, 2012; Carballido et al., 2012; Tschopp et al., 2015) | Stage 1 |
| SMA 0011 | Vertebrae, Skull | Diplodocidae, Galeamopus (Klein and Sander, 2008; Tschopp et al., 2015) | Stage 3 |
| SMA 0014 | Femur | Diplodocidae (this analysis) | Stage 4 |
| SMM P84.15.2 | Femur | Apatosaurus sp. (this analysis) | Stage 1 |
| TMM 993-1 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 4 |
| USNM 2672 | Skull | Diplodocus sp., Diplodocinae Indeterminate (Berman and McIntosh, 1978; Tschopp et al., 2015) | Stage 4 |
| USNM 2673 | Skull | Diplodocus sp.; Galeamopus (Berman and McIntosh, 1978; Tschopp et al., 2015) | Stage 4 |
| USNM 4797 | Femur | Apatosaurus sp. (Wilhite, 2003) | Stage 4 |
| USNM 10865 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 4 |
| USNM 11162 | Skull | Apatosaurus louisae (Berman and McIntosh, 1978) | Stage 4 |
| USNM 337871 | Femur | Diplodocus sp. (Wilhite, 2003) | Stage 1 or 2 |
| WDC BS-157 | Femur | Apatosaurus sp. (Wilhite, 2003) | Stage 3 |
| YPM 429 | Vertebrae | Barosaurus (Lull, 1919; McIntosh, 2005; Tschopp et al., 2015) | Stage 4 |
| YPM 1980 | Vertebrae | Apatosaurus excelsus, Brontosaurus excelsus (Ostrom and McIntosh, 1966; Tschopp et al., 2015) | Stage 4 |
| YPM 5862 | Femur | Apatosaurus sp. (Wilhite, 2003) | Stage 1 |
APPENDIX 3.
The Histo-Morph Ontogeny Scale (H-MOS).
| Stage 1 | Stage 2 | Stage 3 | Stage 4 | |
| HOS | <4 | 5-7 | 8-10 | >10 |
| Postparietal aperture | ? | Present | Present | Absent |
| LAGs (minimum record via dorsal ribs) | <2 | 2-6 | 7 - 15* (15 is an estimated demarcation) |
>15 |
| Cervical pneumatic architecture |
Acamerate with shallow Fossae (4-8 mm) | Procamerate to Camerate with deepening Fossae (7-24 mm) |
Camerate with increasing depth and abundance of Fossae and Foramina | Camerate with extensive and numerous Fossae and Foramina |
| Cervical pneumaticity (CT scan) |
No internal structures | Camerae & Camellae | Thinning median septum with Camerae and Camellae | ? |
| Dorsal pneumaticity (CT scan) |
? | Thinning median septum with Camerae | Thinning median septum with Camerae and Camellae | Extensive Camerae in centrum and arch |
| Ant. cervical bifurcation | No bifurcation | No bifurcation | Notched to weakly bifurcated | Fully bifurcated |
| Post. cervical bifurcation | No bifurcation | Weakly bifurcated | Narrow bifurcation | Fully bifurcated |
| Ant. dorsal bifurcation | No bifurcation | Narrow bifurcation | Narrow bifurcation | Fully bifurcated |
| Post. dorsal bifurcation | No bifurcation | Narrow bifurcation | Bifurcated | Fully bifurcated |
| Ant. Caudal bifurcation | No bifurcation | No bifurcation | No bifurcation | Fully bifurcated |
| Neural spine Trabeculae (mm) |
? | ≤ 1 | ≥ 3 | ? |
| Femoral head orientation | ? | 20° - 11° | 10° - 5° | <5° |
| 4th Trochanter position | Proximal | Proximal | Proximal to mid-diaphysis | Mid-diaphysis |
| Medial Condyle | Not pronounced | Ventrally expanding | Laterally epanding | Greatly pronounced |
| Femur length (mm) | <800 | 800 - 1,200 | 1,200 - 1,450 | >1,450 |
| Body mass (kg) | <2,000 | 2,000 - 3,000 | 3,000 - 6,000 | >6,000 |
APPENDIX 4.
Diplodocidae body mass table using the allometry based body mass formula of Mazzetta (2004) (log Body Mass = 2.955 x log Femur Circumference − 4.166).
| Taxon | Specimen Number | Femur Circumference (mm) | Body Mass (kg) | Mass - Pneumaticity (kg) |
| Apatosaurus sp. | MWC "Moffat Co. Apato." | 908.05 | 49378.59631 | 44440.73667 |
| Apatosaurus | SMA 0014 | 810 | 26827.06928 | 24144.36235 |
| Apatosaurus | AMNH 613 | 805 | 26340.67055 | 23706.60349 |
| Apatosaurus | AMNH 606 | 795 | 25385.44993 | 22846.90494 |
| Apatosaurus | AMNH 7539 | 766 | 22745.52756 | 20470.9748 |
| Apatosaurus | AMNH 353 | 725 | 19332.98035 | 17399.68231 |
| A. louisae | CM 30766 | 701 | 17502.36514 | 15752.12863 |
| Brontosaurus | CM 21785 | 690 | 16703.17623 | 15032.8586 |
| Apatosaurus | CM 85 | 678 | 15859.29017 | 14273.36116 |
| Brontosaurus | CM 87 | 650 | 14000.9972 | 12600.89748 |
| Apatosaurus | CM 566 | 626 | 12527.86759 | 11275.08083 |
| Apatosaurus | CM 33976 | 600 | 11051.89995 | 9946.709959 |
| Apatosaurus | P25112 | 555 | 8777.801694 | 7900.021524 |
| Apatosaurus | OMNH 01667 | 513 | 6956.5965 | 6260.93685 |
| Apatosaurus | KUVP 1351 | 453 | 4816.928356 | 4335.235521 |
| Apatosaurus | BS 157 | 349 | 2228.687292 | 2005.818563 |
| Apatosaurus | CMNH 10039 | 310 | 1570.26809 | 1413.241281 |
| Apatosaurus | USNM 4797 | 288 | 1263.29507 | 1136.965563 |
| Apatosaurus | BYU 601-17103 | 216 | 539.896901 | 485.9072109 |
| Apatosaurus | YPM 5862 | 184.15 | 336.963175 | 303.2668575 |
| Apatosaurus | MWC 5439 | 148 | 176.6539566 | 158.9885609 |
| diplodocid indeterminate | SMM P84.15.2 | 120 | 95.05622917 | 85.55060625 |
| Galeamopus | HMNS 175 | 590 | 10516.4145 | 9464.77305 |
| Diplodocus | USMN 10865 | 564 | 9205.126313 | 8284.613681 |
| Diplodocus | TMM 993-1 | 551 | 8592.172245 | 7732.95502 |
| D. carnegii | CM 94 | 540 | 8095.12502 | 7285.612518 |
| Diplodocus | BYU 725-13643 | 536 | 7919.211712 | 7127.290541 |
| Diplodocus | BYU 725-13369 | 513 | 6956.5965 | 6260.93685 |
| D. carnegii | CM 84 | 510 | 6837.067544 | 6153.36079 |
| Diplodocus | BYU Fe-4-DM197 | 490 | 6074.764083 | 5467.287674 |
| Diplodocus | AMNH 435 | 458 | 4975.737808 | 4478.164027 |
| Diplodocus | BYU 725-11421 | 456 | 4911.804923 | 4420.624431 |
| Diplodocus | AMNH 7539 | 455 | 4880.043319 | 4392.038987 |
| Diplodocus | BYU 725-12155 | 455 | 4880.043319 | 4392.038987 |
| Diplodocus | BB 761 | 451 | 4754.355816 | 4278.920235 |
| Diplodocus | BYU Fe-5-DM172 | 435 | 4273.028154 | 3845.725339 |
| Diplodocus | AMNH 5855 | 410 | 3587.37103 | 3228.633927 |
| Diplodocus | MOR 592-35 | 409 | 3561.577304 | 3205.419574 |
| D. longus | CM 30762 | 387 | 3024.72504 | 2722.252536 |
| Diplodocus | BYU 725-16569 | 385 | 2978.766464 | 2680.889817 |
| Diplodocus | MOR 790 7-23-95-122 | 371.5 | 2680.577713 | 2412.519942 |
| Diplodocus | BYU 725-16610 | 365 | 2544.341575 | 2289.907418 |
| Diplodocus | MOR 790 7-5-95-7 | 362 | 2483.04063 | 2234.736567 |
| Diplodocus | BB 463 | 361 | 2462.826294 | 216.543665 |
| Diplodocus | BYU 725-4889 | 354 | 2324.36667 | 2091.930003 |
| Diplodocus | BYU 725-9026 | 350 | 2247.610597 | 2022.849537 |
| Diplodocus | USMN 337871 | 325 | 1805.574164 | 1625.016747 |
| D. longus | CM 21788 | 317 | 1677.374822 | 1509.63734 |
| D. longus | CM 33991 | 315 | 1646.295103 | 1481.665593 |
| Diplodocus | OMNH 1793 | 254 | 871.5325918 | 784.3793326 |
| Diplodocus | DNM 3781 | 243 | 764.6567709 | 688.1910938 |
| Diplodocus | BYU 725-13670 | 162 | 230.7368014 | 207.6631213 |
| diplodocid indeterminate | P.84.15.2 | 120 | 95.05622917 | 85.55060625 |
APPENDIX 5.
Supplemental Information.
The material contained herein serves to address some of the comments and questions raised by Wedel and Taylor (2013), along with some minor discussion on non-Apatosaurus and Diplodocus genera.
Anterior Cervical Bifurcation. Wedel and Taylor (2013) suggest that there is no evidence of bifurcated neural spines more anteriorly than C6 in any known North American diplodocid. While it is correct that the anterior most neural spines are damaged and reconstructed from Diplodocus carnegii (CM 84), Brontosaurus excelsus (YPM 1980), and Brontosaurus parvus (UWGM 15556), resulting in unknown spine morphology for these specimens, other cervical series from North American diplodocids do show anterior bifurcation (Appendix 6). The anterior-most cervical neural spines from CM 555 (Apatosaurus or Brontosaurus excelsus) are damaged, yet C4 distinctly has weak bifurcation. C5 has weakly expressed bifurcation; while C6 and the remaining vertebrae have fully expressed bifurcation. Even within Barosaurus lentus we see similar anterior-most bifurcation. In AMNH 7530 (alternatively identified as Kaatedocus by Tschopp et al., 2015), the neural spine of C2 is not bifurcated, yet the neural spine of C3 clearly is. After C3, the spines of C4 and C5 are broad, but not bifurcated. The remaining cervical vertebrae are on display, and removal to examine the anterior morphology was not possible at the time of visitation. A slightly larger specimen AMNH 7535 exhibits similar spine morphology. The neural spine of C2 is slightly damaged at the apex, but the neural spine would appear to be bifurcated (if so it would be the anterior most documentation of spine bifurcation). C3 and C5 are missing, and the neural spine of C4 is badly damaged. The neural spine of C6 is clearly bifurcated, yet C7-C13 are un-bifurcated. The final vertebra from this specimen, C14, does have a broad neural spine with incipient or weakly expressed bifurcation.
The claim that no North American diplodocids possessed bifurcated neural spines farther anteriorly than C6 is now shown to be incorrect. Through examination of several complete or nearly complete cervical series, we now see that in diplodocids incipient to weak neural spine bifurcation can occur between C3-C5 (Appendix 6). It is interesting to see that in the anterior most cervical vertebrae there is a small span of bifurcation preceded and followed by a lack of bifurcation. If the hypothesis that bifurcation is developed as a means to maintain and sustain horizontal mobility (Woodruff, 2014) is correct, the presence of select anterior-most bifurcation could be indicative of active cranial mobility. As demonstrated by Wedel and Taylor (2013), serial position is critical. If serial position was unknown in these specimens then there would be a valid argument to place these bifurcated anterior cervicals more posteriorly. From the examined specimens it appears that diplodocids (at least North American taxa) could have neural spine bifurcation prior to C6, and that these damaged historic specimens could be reconstructed with such.
The “End” Of Diplodocid Ontogeny. With the information attained from many of these 2/3 sized diplodocids (such as MOR 592 MOR 7029 [both Diplodocus sp.]), it would appear that once a diplodocid reaches roughly 2/3 its maximum length that proportionally (and potentially mechanically) it is more equivalent to the skeletally mature form. Simply applying force per unit area or a Ponderal Index (Thompson, 1942), the gravitational forces acting upon a 15 meter and 27 meter long animal are more similar (approximately 2 times an increase in both body mass and body length) than those acting upon a 2 meter and a 15 meter long animal (approximately 38 times heavier and 7.5 times longer). Once a diplodocid reaches that 2/3 sized threshold it has achieved the skeletal adaptations needed to support the large vertebral column (i.e., bifurcation of the neural spines). From the 2/3 size through the remainder of its life, the animal then modifies the existing structure to deal with the increasing stresses enacted upon it. This explains why the spine bifurcation changes are less dramatic, and why the centrum begins to expand along with other proportional changes. At the point of dramatic weight increase, it is far easier to modify a pre-existing structure than to suddenly develop a new feature. It would appear that from hatching, an immature diplodocid is in a dramatic ontogenetic race to develop the skeletal features needed to support its eventual gigantic girth.
Woodruff and Fowler (2012) Clarification. In the discussion section Woodruff and Fowler (2012) say, “Just as particularly large diplodocid specimens... have been more recently recognized as large and potentially older individuals of already recognized taxa,... taxa defined on small specimens... might represent immature forms of Diplodocus or Apatosaurus.” Clarifying the original phrasing, the meaning of this passage is that ontogenetic and statigraphic analysis of the characters that diagnose particular taxa within Dipodocoidea (particularly those described based on immature holotypes) may significantly alter the structure of the phylogenetic tree. Further, isolated specimens currently attributed to a given taxon may instead turn out to be ontogenetic stages of a different taxon (without sinking the original designated taxon), these mis-assignments due to heterochronic shifts are only recognizable with stratigraphic and ontogenetic analysis.
Barosaurus. One possible contentious specimen to the ongoing discussion of ontogenetic development of neural spine bifurcation is the immature Barosaurus sp. (DINO 2921) described by Melstrom et al. (2016). While documenting numerous important ontogenetic vertebral characters (ranging from Elongation Index, neurocentral fusion, to pneumatic architecture; and thus further verification of the allometric development of the sauropod skeleton), Melstrom et al. (2016) claim that the morphology of the spine bifurcation in this ~1/3 adult sized individual is indistinguishable from that of a mature animal. The precise spinal morphologies and details are hopefully forthcoming (and respectfully such is not demonstrated nor documented in Melstrom et al. [2016]), but perhaps the specimen DINO 2921 could falsify the hypothesis of Woodruff and Fowler (2012).
Yet we would propose that if Melstrom et al. (2016) are indeed correct about the spine morphology of DINO 2921, this could be an incredibly important key to understanding sauropod evolution within the Morrison Formation. If the stratigraphic resolution is correct (see the informative works of K. Truillo), then DINO 2921 comes from the lower portion of the Brushy Basin Member (Turner and Peterson, 1999; Carpenter, 2013), while other immature Barosaurus specimens (such as AMNH 7530 and 7535; note Tschopp et al. [2015] identify AMNH 7530 as Kaatedocus) are non-bifurcated and come from the upper portion of the Salt Wash Member (Turner and Peterson, 1999; Michelis, 2004; Tschopp and Mateus, 2013). If this resolution, taxonomy, and morphology is correct, then this has substantial implications for Barosaurus heterochrony. Again, all of this hinges on the correct initial identification, but if so, this means that Lower and Upper Morrison specimens have differing vertebral biomechanics (i.e., distinct differing morphologies), and given the stratigraphic and calculated temporal range, this may be initial grounds to begin the examination and inquire into heterochrony, and therefore the possibility of two Morrison Barosaurus taxa.
Camarasaurus. A minor point we would like to bring to light is the recognition of neural spine bifurcation within the basal macronarian Camarasaurus. Woodruff and Fowler (2012) note that in specimens from the Kenton Quarry, the neural spines of immature Camarasaurus sp. are morphologically similar to immature diplodocids in that the neural spines are short and non-bifurcated. No in depth examination on Camarasaurus sp. was carried out in the preliminary analysis (but is currently underway by DCW), but the possibility of Camarasaurus spine ontogeny was favored by Wedel and Taylor (2013). While display features can be modified throughout ontogeny (i.e., male peacock plumage and cassowary and helmeted guinea fowl casques), it would seem unusual that a biomechanical feature could be ontogenetic in one clade, yet static in a closely related clade. In examination of the cervical and dorsal series from the presumed immature Camarasaurus lentus (CM 11338), the neural spines are bifurcated, but the depth of bifurcation is shallow and the neural spine apices are much closer together than in a fully mature animal. A relatively small Camarasaurus sp. specimen at the Great Plains Dinosaur Museum (GPDM 220) has cervical and dorsal neural spines that likewise exhibit shallow bifurcation and narrow neural spines (Woodruff and Foster, 2017). These features alone have been previously thought to be valid autapomorphies of a new genus (N. Murphy and K. Carpenter, personal commun., 2012). However, analysis by Woodruff and Foster (2017) has contrarily demonstrated that GPDM 220 is a maturationally old, small statured individual. Thus GPDM 220 would further verify the complex relationship between vertebral mechanics and ontogeny within sauropods (Woodruff and Foster, 2017).
Haplocanthosaurus. Wedel and Taylor (2013) perform a laudable job verifying that the genus Haplocanthosaurus is not a juvenile Apatosaurus or Diplodocus (the specific lines of reasoning will not be addressed here but we recommend referral to their text; Wedel and Taylor [2013] p. 23-27). Being one of the rarest of Morrison taxa, Haplocanthosaurus is known from three species: H. delfsi, H. priscus, and “H. utterbacki”. Collected from the lower Brushy Basin Member of the Morrison Formation, CM 572 (H. priscus) and CM 879 (“H. utterbacki”) both were found meters away from each other in the Marsh-Felch Quarry 1. While CM 879 has not been histologically sampled to assess maturity, the general consensus (largely based on the overall morphology) is that it represents an immature animal (the differences and validity between H. priscus and H. delfsi shall not be addressed here; McIntosh and Williams [1988], Wedel and Taylor [2013]). As Wedel (2009) illustrates, CM 572 exhibits widespread neurocentral synostosis, whereas CM 879 exhibits primarily completely unfused neural arches (remember from the manuscript that vertebral fusion is not conclusively indicative of maturity within dinosaurs). Though very weakly expressed and exceedingly rare, incipient neural spine bifurcation has been observed within some specimens of Haplocanthosaurus (Appendix 7). Neural spine bifurcation is observed in a posterior dorsal of CM 879, and within an anterior dorsal of CM 572. We are well aware of the importance of serial position and the morphological differences between such vertebrae, however due to the rarity of this feature and the relative proximity a comparison shall still be made. In a posterior cervical of CM 879, the bifurcation is formed by a connection of two closely spaced “humps” (reminiscent of bifurcation observed in CM 555). Within an anterior dorsal of CM 572 these “humps” are spaced and the bifurcation trough is a shallow “V”-shape. Based on the spinal morphology, it would appear that, unlike other members of Diplodocidae, Haplocanthosaurus did not biomechanically require neural spine bifurcation. In conjunction with McIntosh and Williams (1988) and Wedel and Taylor (2013), we would consider “H. utterbacki” as a nomen dubium, and further agree that it represents an immature form of H. priscus. In addition to this, the variation between the two specimens would suggest that while incipient, the neural spine bifurcation observed in Haplocanthosaurus may also be ontogenetic. Verification of this point requires further specimens and overlapping material.
In regards to the phylogenetic assignment of Haplocanthosaurus, we would again stress the need for the recognition of ontogenetic stages. In his analysis of Diplodocoidea, J. Whitlock (2011a) shows that the character matrix for Haplocanthosaurus is a combination of several specimens including CM 879 (“H. utterbacki”), CM 572 (H. priscus), and CMNH 10380 (H. delfsi). While these individuals were included to complete otherwise missing characters from other incomplete specimens, the taxonomic uncertainty of Haplocanthosaurus could be due to the fact that the characters states representing it are from a combination of varying ontogenetic stages and potentially separate species (sensu Mannion et al., 2012).
APPENDIX 6.
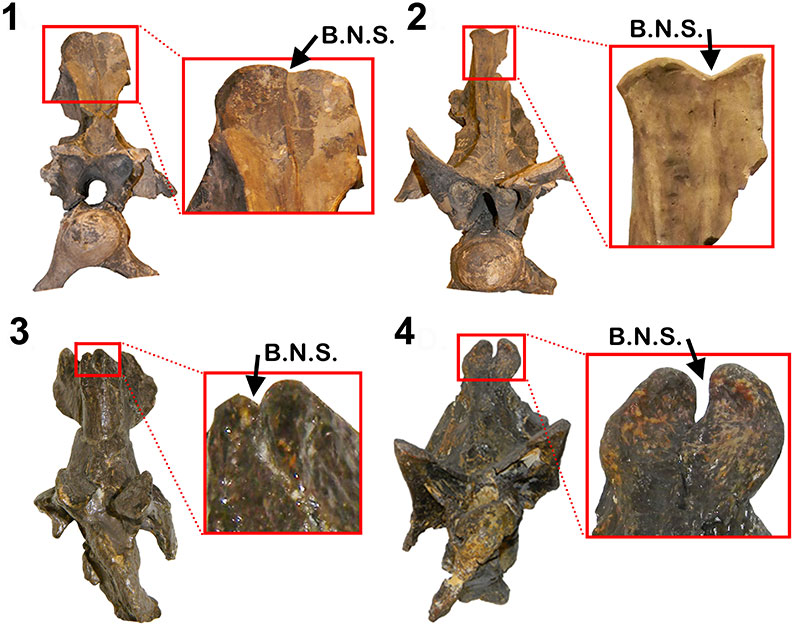
Neural spine bifurcation in diplodocid anterior most cervical vertebrae. 1-2, Apatosaurus CM 555; 3,Barosaurus AMNH 7530; 4,Diplodocus sp. MOR 592. B.N.S = Bifurcated Neural Spine. All cervical vertebrae in anterior view. Not to scale. Scale bars equal 10 cm.
APPENDIX 7.
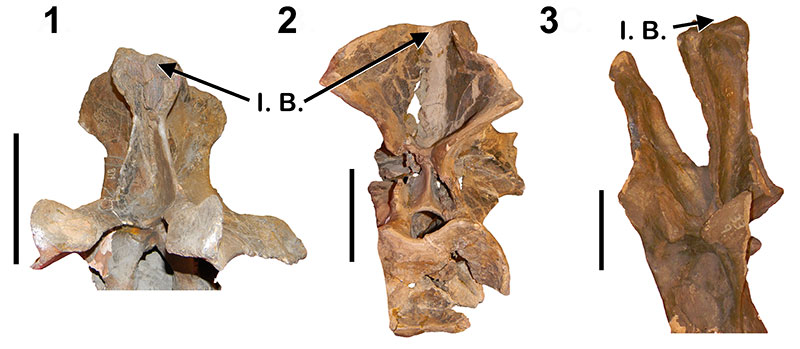
Haplocanthosaurus incipient neural spine bifurcation. 1, Haplocanthosaurus “utterbacki ” (CM 879) anterior dorsal in anterior view; 2,Hapalocanthosauruspriscus (CM 529) anterior dorsal in posterior view; 3,H. “utterbacki ” (CM 879) posterior dorsal in anterior view. I.B. = Incipient Bifurcation. Not to scale. Scale bar equals 10 cm.
APPENDIX 8.
Elongation Index of diplodocid cervical vertebrae (EI = centrum length divided by cotyle diameter).
| Diplodocus | |||
| HQ 1 SMA 0003 | |||
| Centrum Length (mm) | Cotyle Diameter (mm) | EI | |
| C2 | 84 | 37 | 2.27027 |
| C3 | 105.5 | 31 | 3.40323 |
| C4 | 131 | 46.5 | 2.8172 |
| C5 | 184 | 30.5 | 6.03279 |
| C6 | 228 | 46 | 4.95652 |
| C7 | 280 | 44 | 6.36364 |
| C8 | 376 | 55.5 | 6.77477 |
| C9 | 375 | 71 | 5.28169 |
| C10 | 385 | 49 | 7.85714 |
| C11 | 431 | 75 | 5.74667 |
| C12 | 458 | 74 | 6.18919 |
| C13 | 454 | 58.5 | 7.76068 |
| C14 | 463 | 77 | 6.01299 |
| C15 | 474 | 77 | 6.15584 |
| HQ 2 SMA 0004 | |||
| Centrum Length (mm) | Cotyle Diameter (mm) | EI | |
| C2 | 83.5 | 35 | 2.38571 |
| C3 | 107 | 27 | 3.96296 |
| C4 | 132 | - | - |
| C5 | 158 | 28 | 5.64286 |
| C6 | 192 | 26.5 | 7.24528 |
| C7 | 218 | 37 | 5.89189 |
| C8 | 247 | 34.5 | 7.15942 |
| C9 | 264 | 38 | 6.94737 |
| C10 | 296 | 53 | 5.58491 |
| C11 | 295 | 51 | 5.78431 |
| C12 | 316 | 56 | 5.64286 |
| C13 | 326 | 66 | 4.93939 |
| C14 | 314 | 61 | 5.14754 |
| MOR 592 | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C2 | 114.4 | 41.2 | 2.7767 |
| C3 | 128.3 | 41.9 | 3.06205 |
| C4 | - | - | - |
| C5 | 163.6 | 49 | 3.33878 |
| C6 | - | - | - |
| C7 | 247 | 37.5 | 6.58667 |
| C8 | 256.5 | 47.4 | 5.41139 |
| C9 | 271.6 | 67.8 | 4.0059 |
| C10 | 291 | 62.4 | 4.66346 |
| C11 | 279.4 | 103.5 | 2.69952 |
| C12 | 266.7 | 112.5 | 2.37067 |
| C13 | 239.2 | 180.8 | 1.32301 |
| C14 | 349.3 | 73.2 | 4.77186 |
| C15 | 304.8 | 96.2 | 3.1684 |
| CM 84 | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C2 | 165 | 54 | 3.05556 |
| C3 | 243 | 69 | 3.52174 |
| C4 | 289 | 81 | 3.5679 |
| C5 | 372 | 94 | 3.95745 |
| C6 | 442 | 99 | 4.46465 |
| C7 | 485 | 114 | 4.25439 |
| C8 | 512 | 120 | 4.26667 |
| C9 | 525 | 159 | 3.30189 |
| C10 | 595 | 175 | 3.4 |
| C11 | 605 | 210 | 2.88095 |
| C12 | 627 | 225 | 2.78667 |
| C13 | 638 | 231 | 2.7619 |
| C14 | 642 | 295 | 2.17627 |
| C15 | 595 | 245 | 2.42857 |
| Apatosaurus | |||
| CM 555 | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C2 | 140 | 63 | 2.22222 |
| C3 | 221 | 84 | 2.63095 |
| C4 | 274 | 96 | 2.85417 |
| C5 | 270 | 60 | 4.5 |
| C6 | 295 | 114 | 2.58772 |
| C7 | 316 | - | - |
| C8 | 344 | 113 | 3.04425 |
| C9 | 380 | - | - |
| C10 | - | - | - |
| C11 | 480 | 193 | 2.48705 |
| C12 | 460 | 245 | 1.87755 |
| C13 | - | - | - |
| C14 | 378 | 260 | 1.45385 |
| C15 | - | - | - |
| CM 563 | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C3 | 250 | 80 | 3.125 |
| C4 | 300 | 125 | 2.4 |
| C5 | 342 | 134 | 2.55224 |
| C6 | - | - | - |
| C7 | 415 | 170 | 2.44118 |
| C8 | 415 | 205 | 2.02439 |
| C9 | 445 | 215 | 2.06977 |
| C10 | 475 | 250 | 1.9 |
| NSMT-PV 20375 | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C3 | 352 | 97 | 3.62887 |
| C4 | - | - | - |
| C5 | 375 | 155 | 2.41935 |
| C6 | 395 | 195 | 2.02564 |
| C7 | 420 | 220 | 1.90909 |
| C8 | 395 | 195 | 2.02564 |
| C9 | 380 | 235 | 1.61702 |
| C10 | 390 | - | - |
| C11 | - | - | - |
| C12 | 475 | 240 | 1.97917 |
| C13 | - | - | - |
| C14 | 450 | 305 | 1.47541 |
| CM 3018 | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C2 | 190 | 85 | 2.23529 |
| C3 | 280 | 100 | 2.8 |
| C4 | 370 | 100 | 3.7 |
| C5 | - | - | - |
| C6 | 440 | 150 | 2.93333 |
| C7 | 450 | 190 | 2.36842 |
| C8 | 485 | 225 | 2.15556 |
| C9 | 510 | 230 | 2.21739 |
| C10 | 530 | 250 | 2.12 |
| C11 | 550 | 240 | 2.29167 |
| C12 | 490 | 265 | 1.84906 |
| C13 | 480 | - | - |
| Barosaurus | |||
| AMNH 7530 | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C2 | 80 | 14 | 5.71429 |
| C3 | 75 | 26 | 2.88462 |
| C4 | 97 | 37 | 2.62162 |
| C5 | 123 | 62 | 1.98387 |
| C6-C12 are on display and could not be measured first hand | |||
| AMNH 7535 | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C2 | 87 | 15 | 5.8 |
| C3 | - | - | - |
| C4 | 102.5 | 23 | 4.45652 |
| C5 | - | - | - |
| C6 | 135 | 25 | 5.4 |
| C7 | 140 | 21 | 6.66667 |
| C8 | 166 | 36 | 4.61111 |
| C9 | - | - | - |
| C10 | - | - | - |
| C11 | 234 | 32 | 7.3125 |
| C12 | - | - | - |
| C13 | 281 | 30 | 9.36667 |
| C14 | 323 | 47 | 6.87234 |
| AMNH 6341 (from McIntosh, 2005) | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C8 | 618 | 130 | 4.75385 |
| C9 | 685 | 123 | 5.56911 |
| C10 | 737 | 168 | 4.3869 |
| C11 | 775 | 145 | 5.34483 |
| C12 | 813 | 155 | 5.24516 |
| C13 | 850 | 180 | 4.72222 |
| C14 | 865 | 155 | 5.58065 |
| C15 | 840 | 160 | 5.25 |
| C16 | 750 | 250 | 3 |
| YPM 429 (from McIntosh, 2005) | |||
| Centrum Length (mm) |
Cotyle Diameter (mm) |
EI | |
| C13 | 930 | 220 | 4.22727 |
| C14 | 890 | 345 | 2.57971 |
| C15 | - | 300 | - |
| C16 | 720 | 365 | 1.9726 |
