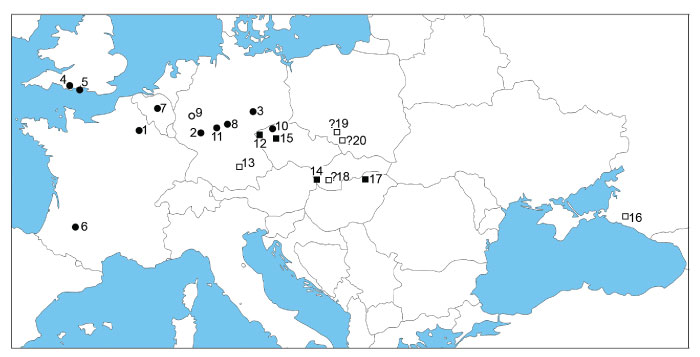
FIGURE 1. Eopelobates aff. E. bayeri from Volchaya Balka locality, Russia (late Miocene, early Turolian). 1-2, frontoparietal (GIN 1143-200) in dorsal (1) and ventral (2) views; 3-4, frontoparietal (GIN 1143-201) in dorsal (3) and ventral (4) views; 5-6, right maxilla (GIN 1143-202) in labial (5) and lingual (6) views; 7-8, right maxilla (GIN 1143-203) in labial (7) and lingual (8) views; and 9-10, ?left maxilla (GIN 1143-204) in labial (9) and lingual (10) views. Scales equal 1 mm.
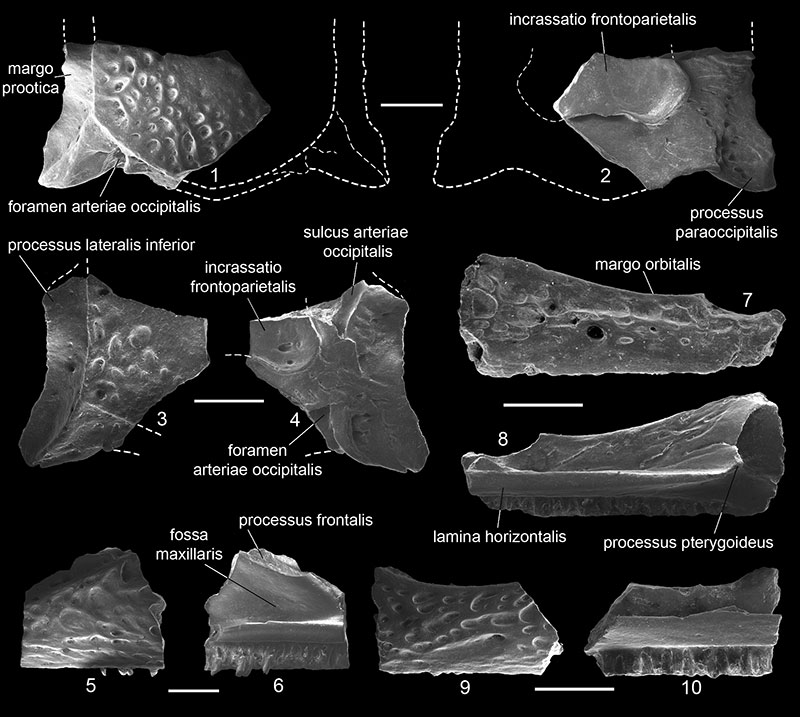
FIGURE 2. Eopelobates aff. E. bayeri from Volchaya Balka locality, Russia (late Miocene, early Turolian). 1-3, presacral vertebra (GIN 1143-212) in dorsal (1), ventral (2), and lateral (3) views; 4-5, presacral vertebra (GIN 1143-213) in dorsal (4) and lateral (5) views; 6-8, sacral vertebra (GIN 1143-221) in dorsal (6), ventral (7), and anterior (8) views; and 9-11, left ilium (GIN 1143-222) in lateral (9) and medial (10) views, and outline of the junctura ilioischiadica in caudal view (11). Scales equal 1 mm.
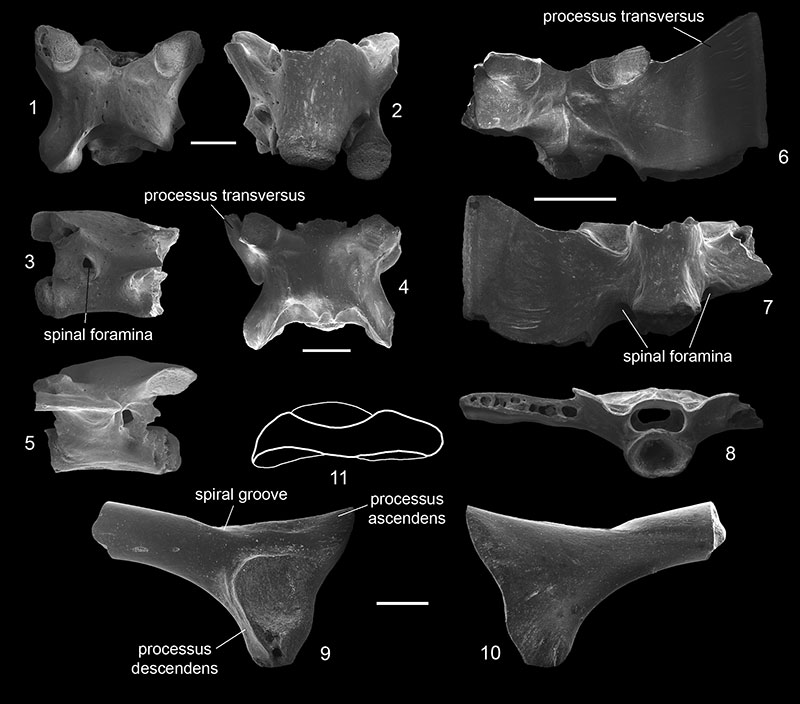
FIGURE 3. Pelobates sp. (1-11) and Pelobatidae indet. (12-15) from Volchaya Balka and Gaverdovsky localities, Russia (late Miocene, early Turolian). 1-2, frontoparietal (GIN 1143-223) in dorsal (1) and ventral (2) views; 3-4, left maxilla (GIN 1143-225) in labial (3) and lingual (4) views; 5-6, right maxilla (GIN 1143-226) in labial (5) and lingual (6) views; 7-9, presacral vertebra (GIN 1143-231) in dorsal (7), ventral (8), and lateral (9) views; 10-11, sacral vertebra (GIN 1144-201) in dorsal (10) and ventral (11) views; 12-13, premaxilla (GIN 1144-202) in lingual (12) and labial (13) views; and 14-15, left scapula (GIN 1143-233) in dorsal (14) and ventral (15) views. Scales equal 1 mm.
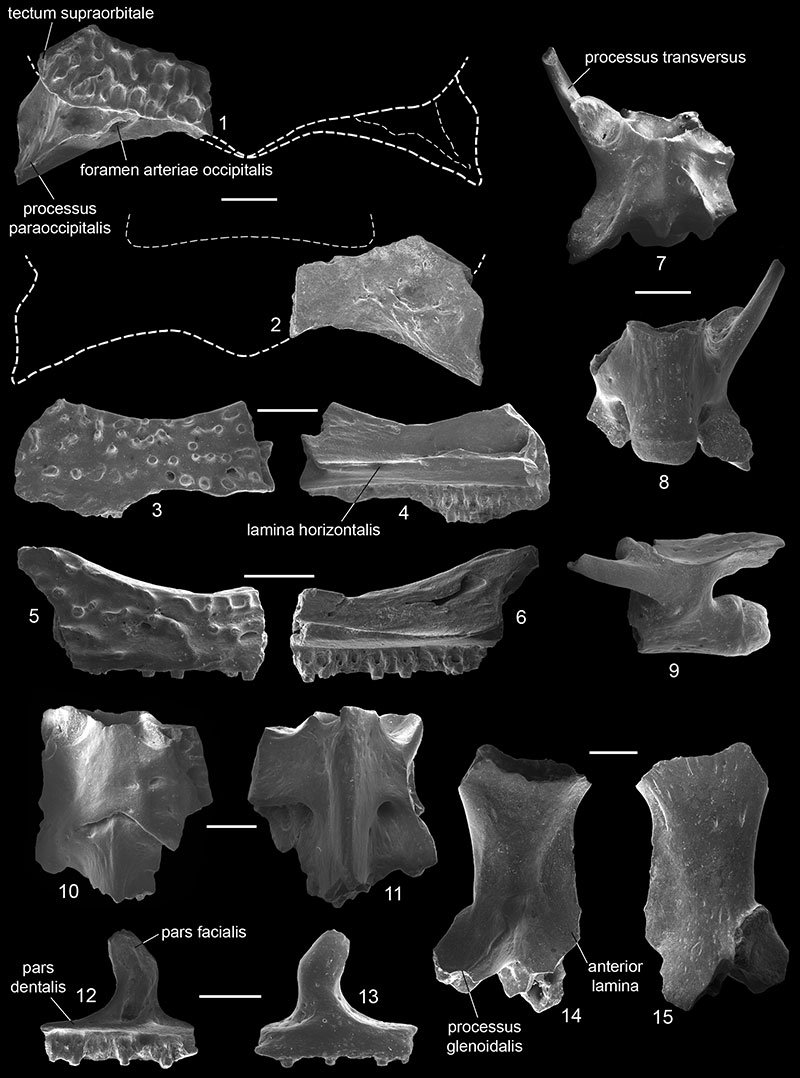
FIGURE 4. Occurrences of Eopelobates from the Paleogene (circles) and Neogene (squares) of Europe. Co-occurrences with Pelobates are indicated by open symbols. 1, Prémontré in France (Eopelobates aff. E. hinschei), MP 10, early Eocene (Duffaud, 2000); 2, Messel in Germany (E. wagneri), MP 11, middle Eocene (Wuttke, 2012); 3, Geiseltal in Germany (E. hinschei), MP 13, middle Eocene (Estes, 1970); 4, Hordle Cliff in UK (Eopelobates cf. E. hinschei), late Eocene (Milner et al., 1982); 5, Headon Hill in UK (cf. Eopelobates), late Eocene (Rage and Ford, 1980); 6, Quercy in France (cf. Eopelobates), late Eocene (Crochet et al., 1981); 7, Hoogbutsel, Hoeleden, and Boutersem TGV in Belgium (E. bayeri), MP 21, early Oligocene (Smith, 2003); 8, Sieblos in Germany (Eopelobates sp.), early Oligocene (Gaudant, 1985); 9, Rott in Germany (E. anthracinus), MP 30, late Oligocene (Parker, 1929); 10, Bechlejovice in Czech Republic (E. bayeri), late Oligocene (Špinar, 1952, 1972); 11, Oberleichtersbach in Germany (Eopelobates sp.), MP 30, late Oligocene (Böhme, 2008); 12, Dolnice in Czech Republic (Eopelobates sp.), MN 4, early Miocene (Hodrová, 1987b); 13, Sandelzhausen in Germany (Eopelobates sp.), MN 5, early Miocene (Böhme, 2010); 14, Devínska Nová Ves in Slovakia (E. bayeri), middle Miocene (Hodrová, 1988); 15, Suchomasty in Czech Republic (Eopelobates sp.), MN 10, late Miocene (Hodrová, 1987a); 16, Volchaya Balka in Russia (Eopelobates aff. E. bayeri), MN 11, late Miocene (this paper); 17, Osztramos 1 in Hungary (Eopelobates sp.), MN 14, Pliocene (Venczel, 2001); 18, ?Ivanovce in Slovakia (? Eopelobates cf. bayeri), MN 15, Pliocene (Hodrová, 1981); 19, ?Węże 1 in Poland (Eopelobates sp.), MN 15, Pliocene (Młynarski, 1961, 1962; Sanchíz and Mlynarski, 1979); and 20, ?Rębielice Królewskie 1 in Poland (Eopelobates sp.), MN16, Pliocene (Sanchíz and Mlynarski, 1979). Data on taxonomic composition of Eopelobates are based mainly on Roček et al. (2014). The unconfirmed occurrences are denoted by a question-mark.