

APPENDICES
APPENDIX 1. Information on the hipparionin sample.
APPENDIX 1.1. Western Mediterranean
The Vallesian hipparionins from the western Mediterranean area come from the fossil sites of Can Llobateres, Can Poncic and Santiga. All three localities are situated on the Vallès-Penedès Basin (northeastern Iberian Peninsula) (Figure 1, main text) and are placed on medial to distal facies of alluvial origin (Casanovas-Vilar et al., 2014). The classical fossil site of Can Llobateres registers the early/late Vallesian (MN9/MN10) transition and has been divided into two different levels with estimated ages of 9.72 Ma and 9.65 Ma, respectively (Casanovas-Vilar et al., 2011). Both Can Poncic and Santiga were dated on the early Vallesian with an estimated age of 10.4 to 9.9 Ma (Casanovas-Vilar et al., 2011). Based on a Vallesian hipparionin cranium of the Vallès-Penedès, Pirlot (1956) established the species Hipparion catalaunicum. Thereafter, Vallesian hipparionins from this basin have been generally assigned to this taxon. The similarities between this form with the central European Hipparion primigenium [= Hippotherium primigenium sensu Bernor et al. (1996)] (Forsten, 1968; Woodburne and Bernor, 1980) have led some authors to include it within the H. primigenium species (Forsten, 1968; Alberdi, 1974; Tomàs et al., 2010), and differentiate it at subspecific level (Alberdi, 1974; Tomàs et al., 2010). The presence of one single large species have been proposed in Can Poncic and Can Llobateres (Scott et al., 2013). We consider both populations to pertain to the H. primigenium catalaunicum subspecies following Tomàs et al. (2010). In Santiga, however, it is evident that at least two species are present, an abundant medium-sized to small hipparionin and a much rarer large one (Forsten, 1997). Although unnamed, the small/medium-sized hipparionin from Santiga represent a different taxon compared to larger Vallesian forms, although it might also be related to the primigenium group. Here we refer to this form as Hipparion sp. (? primigenium small). Due to the lack of dentognathic material of the large species from Santiga, we only analyze the microwear texture of the small hipparionin from this site.
The Turolian material from this area come from the localities of Concud, El Arquillo and Venta del Moro. The classical fossil sites from Concud and El Arquillo are located in the Teruel Basin (central eastern Iberian Peninsula), and Venta del Moro in the Cabriel Basin (central eastern Iberian Peninsula) (Figure 1, main text). Both Concud and Venta del Moro sites were formed in marginal lacustrine environments with abundant vegetation (Montoya et al., 2006; Pesquero et al., 2013). The locality of Concud (MN12) has an estimated age of 7.2-7.1 Ma, while El Arquillo and Venta del Moro are dated on the MN13 with estimated ages of 6.2 Ma (van Dam et al., 2001) and 5.8-5.5 Ma (Opdyke et al., 1997), respectively. Recently, however, a magnetostratigraphic study by Gibert et al. 2013 dated Venta del Moro at 6.23 Ma. Still, El Arquillo site is deemed to be older because of the absence of Paraethomys miocaenicus, similarly to Las Casiones, which was dated at 6.33 Ma (van Dam, 1997; van Dam et al., 2006). During the middle Turolian (MN12) the most common species was Hipparion concudense, a medium to large sized hipparionin [178 kg (Pesquero and Alberdi, 2012)] that is very abundant in Concud (Pesquero and Alberdi, 2012). On the other hand, at least three different sized hipparionins inhabited the Teruel Basin during the late Turolian (MN13). The three sympatric taxa identified in El Arquillo have been referred to Hipparion periafricanum, Hipparion gromovae, and H. primigenium truyolsi (Alberdi, 1974). The two dwarf hipparionins of this site, H. periafricanum and H. gromovae, are the smallest members of this clade in the Iberian Peninsula (Ortiz-Jaureguizar and Alberdi, 2003; Pesquero, 2003). H. primigenium truyolsi, on the other hand, was much larger (Ortiz-Jaureguizar and Alberdi, 2003). An association of three sympatric forms has also been reported in the Venta del Moro. Morphometric comparisons of the most common hipparionin from this site has allowed its assignment to the species Hipparion matthewi (Pesquero et al., 2007), which is also present in eastern Mediterranean basins. This identification, however, has recently been questioned (Koufos, 2016b; Koufos and Vlachou, 2016), so here we decided to refer this medium-sized hipparionin as H. cf. matthewi and to differentiate it from the eastern Mediterranean forms. Pesquero et al. (2007), based on the material from Venta del Moro, also proposed the synonymy of H. gromovae and H. matthewi. Because we are interested in testing dietary differences related to size, the body mass differences between H. gromovae from El Arquillo (84 kg) and H. cf. matthewi from Venta del Moro (123 kg) (Table 1, main text), together with the lack of cranial traits to compare, make us consider both populations two different sets of data. Besides H. cf. matthewi, there are two other hipparionins present in Venta del Moro: a much rarer large form determined as H. primigenium, and a probable smaller subspecies of H. cf. matthewi (Pesquero et al., 2007). Due to the scarcity of remains assigned to these two last forms, here we only analyze the microwear texture of the medium-sized H. cf. matthewi from Venta del Moro. All the hipparionin teeth and mandibles from western Mediterranean sites are stored at the collections of the Institut Català de Paleontologia Miquel Crusafont (ICP), Barcelona, and of the Museo Nacional de Ciencias Naturales (MNCN), Madrid.
APPENDIX 1.2. Balkans
The Vallesian hipparionins pertain to the localities of Ravin de la Pluie and Pentalophos from the Axios Valley (southeastern Balkan Peninsula), and to Nikiti-1, which is located in the Chalkidiki Peninsula (southeastern Balkan Peninsula) (Figure 1, main text). Ravin de la Pluie was dated to the lower part of the late Vallesian (MN10) with an estimated age of ~9.3 Ma based on a combination of biochronological and paleomagnetostratigraphical data (Sen et al., 2000; Koufos, 2006a). Nikiti-1 is correlated based on its mammalian assemblage to the upper part of the late Vallesian with an estimated age ranging from 9.3 to 8.7 Ma (Koufos, 2006b, 2016a). Pentalophos’ chronology is not clear, although its faunal association indicates that it could be slightly older than Ravin de la Pluie (Koufos, 2006a, 2006b, 2013). Following Vlachou (2013), two hipparionin morphotypes were present in Macedonia during the Vallesian: the small macedonicum and the large primigenium morphotype. The material of the small macedonicum morphotype has been assigned to the Hipparion macedonicum species, while the larger hipparionins of the primigenium morphotype have been referred to Hipparion aff. giganteum in Nikiti-1, and to Hipparion cf. sebastopolitanum in Pentalophos and Ravin de la Pluie (Koufos et al., 2016; Vlachou, 2013).
Numerous Turolian localities have yielded rich hipparionin assemblages in the Balkan region. The Turolian sample analyzed here comes from the localities of Nikiti-2, Ravin des Zouaves-5, Hadjidimovo, Perivolaki and Dytiko. The oldest fossil site is Nikiti-2 (Chalkidiki Peninsula) (Figure 1, main text), dated to the early Turolian (MN11) with an age comprised between 8.7-8.2 Ma (Koufos, 2016a). Four different hipparionins are recognized here: two small taxa of the macedonicum morphotype (Hipparion sithonis and H. macedonicum), a larger form that pertains to the dietrichi morphotype (Hipparion philippus), and a rare and large hipparionin of the proboscideum group identified as Hipparion cf. proboscideum (Vlachou, 2013; Koufos and Vlachou, 2016). Ravin des Zouaves-5 (Axios Valley) is also an early Turolian site (MN11) dated at ~8.2 Ma (Koufos, 2006b). In this locality, Koufos and Vlachou (2019) reported five different taxa: H. macedonicum and H. sithonis of the macedonicum morphotype, H. proboscideum and H. cf. mediterraneum of the proboscideum morphotype, and H. philippus of the dietrichi morphotype. From this site, here we analyzed mainly specimens of H. macedonicum due to their availability. Hadjidimovo locality (Mesta Valley, central Balkan Peninsula) (Figure 1, main text) was dated between the early-middle Turolian (MN11-MN12) with an estimated age of ~7.5 Ma (Spassov, 2002). Three species have been identified, although the major bulk of the material corresponds to the two large species Hipparion mediterraneum [ proboscideum morphotype sensu Vlachou (2013)] and Hipparion brachypus [ primigenium morphotype sensu Vlachou (2013)] (Hristova, 2009), which are the species herein analyzed. We also sampled the hipparionins from Perivolaki (Thessaly, southeastern Balkan Peninsula) (Figure 1, main text), a site dated on the upper middle Turolian (MN12) with an estimated age of 7.3-7.1 Ma (Koufos, 2006a, 2006b). In Perivolaki, three different species are found: H. macedonicum, H. philippus and H. proboscideum; forming a similar association than other early and middle Turolian sites from the Axios Valley. Finally, the youngest locality is represented by the three Dytiko sites (DKO, DTK, DIT; Axios Valley) (Figure 1, main text), which were dated to the late Turolian (MN13) (Koufos, 2006b). Three hipparionin taxa are identified in the Dytiko faunal assemblage: the large Hipparion aff. platygenys [ dietrichi morphotype sensu Vlachou (2013)], the small Hipparion cf. macedonicum, and a medium-sized rare form resembling to Hipparion moldavicum and tentatively assigned to the proboscideum morphotype (Vlachou, 2013). Because of the scarcity of some of the material from the species mentioned, we did not consider for analysis the proboscideum group from Nikiti-2 nor from the Dytiko sites, and only include one specimen from Ravin des Zouaves-5. The fossil material analyzed here from Hadjidimovo is stored at the Asenovgrad Museum (a division of the National Museum of Natural History of Sofia, Bulgaria) and the specimens from the Greek localities come from the Laboratory of Geology and Paleontology, Aristotle University of Thessaloniki, Greece.
APPENDIX 1.3. Western Anatolia
The Turolian hipparionins from the western Anatolian bioprovince come from the localities of Şerefköy-2, Gülpınar and the Samos new collections. The specimens from Samos pertain to the MTLA/B sites found in the Adrianos ravine during the recent excavations (Mytilinii Basin, Samos Island) (Figure 1, main text), which were dated on the late middle Turolian (~7.1 Ma) (Koufos et al., 2009; Vlachou and Koufos, 2009). At least five hipparionin taxa pertaining to four different morphotypes are present there: H. cf. matthewi (macedonicum morphotype), Hipparion aff. forstenae (proboscideum morphotype), H. cf. proboscideum (proboscideum morphotype), H. dietrichi (dietrichi morphotype) and H. brachypus (primigenium morphotype) (Vlachou and Koufos, 2009; Vlachou, 2013). In the Anatolian Peninsula, the locality of Şerefköy-2 (Muğla Yatağan Basin, western Anatolia) (Figure 1, main text) has been referred to the middle Turolian (MN12) based on its fossil mammal assemblage (Kaya et al., 2012). Four to five different hipparionin forms might be present in this site, a rare and large species similar to H. brachypus, two medium hipparionins broadly referred as Hipparion, and two small forms only identified as belonging to the Cremohipparion group [sensu Bernor et al. (1996)] (Kaya et al., 2012). The medium-sized hipparionins show similarities to H. mediterraneum specimens (Kaya et al., 2012). In consequence, we included here the medium sized forms to the proboscideum group. We further relate the small hipparionins of Şerefköy-2 to the species H. matthewi, as it has been done with the majority of the small-sized forms from eastern Mediterranean basins (Koufos, 2016b), thus including them to the macedonicum group defined by Vlachou (2013). Finally, Gülpınar fossil site is located on the Biga Peninsula (northwestern Anatolia) (Figure 1, main text) and corresponds to a sedimentary sequence of fluvial origin dated on early-middle Turolian (MN11-MN12) (Forsten and Kaya, 1995; Koufos et al., 2018). Three different forms have been distinguished in Gülpınar. The smallest hipparionin has been identified as H. cf. matthewi (macedonicum morphotype) and the medium-sized form show affinities to H. dietrichi from the classic Samos quarries (Forsten and Kaya, 1995). Due to these similarities, here we include this form to the dietrichi group defined by Vlachou (2013). The larger hipparionin remains from Gülpınar have not been determined. We did not analyze the dental microwear textures of the largest taxon--usually related to H. brachypus (Vlachou and Koufos, 2009; Kaya et al., 2012)--from the western Anatolian fossil sites due to its scarcity. The material from Samos is stored at the Paleontological Museum of Mytilinii, Samos, Greece, and the material from continental Anatolian sites at the Natural History Museum of Ege University of Izmir, Turkey.
APPENDIX 1.4. References
Alberdi, M.T. 1974. Las “faunas de Hipparion ” de los yacimientos españoles. Estudios Geológicos, 30:189-212.
Bernor, R.L., Koufos, G.D., Woodburne, M.O., and Fortelius, M. 1996. The evolutionary history and biochronology of European and Southwest Asian Late Miocene and Pliocene hipparionine horses, p. 307-338. In Bernor, R.L., Fahlbusch, V., and Mittmann, H.-W. (eds.), The Evolution of Western Eurasian Neogene Mammal Faunas. Columbia University Press, New York.
Casanovas-Vilar, I., Alba, D.M., Garcés, M., Robles, J.M., and Moyà-Solà, S. 2011. Updated chronology for the Miocene hominoid radiation in Western Eurasia. Proceedings of the National Academy of Sciences of the United States of America, 108:5554-9. https://doi.org/10.1073/pnas.1018562108
Casanovas-Vilar, I., Van den Hoek Ostende, L.W., Furió, M., and Madern, P.A. 2014. The range and extent of the Vallesian Crisis (Late Miocene): New prospects based on the micromammal record from the Vallès-Penedès basin (Catalonia, Spain). Journal of Iberian Geology, 40:29-48. https://doi.org/10.5209/rev_JIGE.2014.v40.n1.44086
Forsten, A. 1968. Revision of the palearctic Hipparion. Acta Zoologica Fennica, 119:1-134.
Forsten, A. 1997. Hipparion from Santiga (Spain) and its biostratigraphic significance. Paleontologia i Evolució, 30-31:77-82.
Forsten, A. and Kaya, T. 1995. The hipparions (Mammalia, Equidae) from Gülpinar (Canakkale, Turkey). Paläontologische Zeitschrift, 69:491-501. https://doi.org/10.1007/BF02987809
Gibert, L., Scott, R., G., Montoya, P., Ruiz-Sánchez, J.F., Morales, J., Luque, L., Abella, J. and Lería, M. 2013. Evidence for an African-Iberian mammal dispersal during the pre-evaporitic Messinian. Geology, 41:691-694. https://doi.org/10.1130/g34164.1
Hristova, L. 2009. Ontogeny and variability in the cheek region of hipparions from the late Miocene locality of Hadzhidimovo-1, southwest Bulgaria. Rivista Italiana di Paleontologia e Stratigrafia, 115:125-132. https://doi.org/10.13130/2039-4942/5924
Kaya, T., Mayda, S., Kostopoulos, D.S., Alcicek, M.C., Merceron, G., Tan, A., Karakutuk, S., Giesler, A.K., and Scott, R.S. 2012. Şerefköy-2, a new Late Miocene mammal locality from the Yataĝan Formation, Muĝla, SW Turkey. Comptes Rendus - Palevol,11:5-12. https://doi.org/10.1016/j.crpv.2011.09.001
Koufos, G.D. 2006a. Palaeoecology and chronology of the Vallesian (late Miocene) in the Eastern Mediterranean region. Palaeogeography, Palaeoclimatology, Palaeoecology, 234:127-145. https://doi.org/10.1016/j.palaeo.2005.01.014
Koufos, G.D. 2006b. The Neogene mammal localities of Greece: Faunas, chronology and biostratigraphy. Hellenic Journal of Geosciences, 41:183-214.
Koufos, G.D. 2013. Neogene mammal biostratigraphy and chronology of Greece, p. 595-621. In Wang, X., Flynn, L.J., and Fortelius, M. (eds.), Fossil Mammals of Asia--Neogene Biostratigraphy and Chronology. Columbia University Press, New York.
Koufos, G.D. 2016a. History, stratigraphy and fossiliferous sites, p. 3-10. In Koufos G.D. and Kostopoulos, D.S. (eds.), Palaeontology of the Upper Miocene Vertebrate Localities of Nikiti (Chalkidiki Peninsula, Macedonia, Greece).Geobios, 49. https://doi.org/10.1016/j.geobios.2016.01
Koufos, G.D. 2016b. Hipparion macedonicum revisited: New data on evolution of hipparionine horses from the Late Miocene of Greece. Acta Palaeontologica Polonica, 61:519-536. https://doi.org/10.4202/app.00169.2015
Koufos, G.D., Kostopoulos, D.S., and Merceron, G. 2009. Palaeoecology - Palaeobiogeography, p. 409-430. In Koufos, G.D. and Nagel, D. (eds.), The late Miocene Mammal Faunas of the Mytilinii Basin, Samos Island, Greece: new collection. Beiträge zur Paläontologie, 31.
Koufos, G.D., Kostopoulos, D.S., and Vlachou, T.D. 2016. Revision of the Nikiti 1 (NKT) fauna with description of new material, p. 11-22. In Koufos G.D. and Kostopoulos, D.S. (eds.), Palaeontology of the Upper Miocene Vertebrate Localities of Nikiti (Chalkidiki Peninsula, Macedonia, Greece).Geobios, 49. https://doi.org/10.1016/j.geobios.2016.01.006
Koufos, G.D., Mayda, S., and Kaya, T. 2018. New carnivoran remains from the Late Miocene of Turkey. Paläontologische Zeitschrift, 92:131-162. https://doi.org/10.1007/s12542-017-0376-2
Koufos, G.D., Vlachou, T.D., 2016. Equidae, p. 85-118. In Koufos G.D. and Kostopoulos, D.S. (eds.), Palaeontology of the Upper Miocene Vertebrate Localities of Nikiti (Chalkidiki Peninsula, Macedonia, Greece).Geobios, 49. https://doi.org/10.1016/j.geobios.2016.01.001
Koufos, G.D. and Vlachou, T.D. 2019. New material and revision of hipparions from the Upper Miocene locality Ravin des Zouaves-5, RZO of Axios Valley (Macedonia, Greece). Geobios, 57:33-52. https://doi.org/10.1016/j.geobios.2019.10.004
Montoya, P., Morales, J., Robles, F., Abella, J., Benavent, J. V, Marín, M. D., and Ruiz Sánchez, F.J. 2006. Las nuevas excavaciones (1995-2006) en el yacimiento del Mioceno final de Venta del Moro, Valencia. Estudios Geológicos, 62:313-326.
Opdyke, N., Mein, P., Lindsay, E., Perez-Gonzales, A., Moissenet, E., and Norton, V.L. 1997. Continental deposits, magnetostratigraphy and vertebrate paleontology, late Neogene of eastern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology, 133:129-148. https://doi.org/10.1016/S0031-0182(97)00080-1
Ortiz-Jaureguizar, E. and Alberdi, M.T. 2003. El patrón de cambios en la masa corporal de los Hipparionini (Perissodactyla, Equidae) de la Península Ibérica durante el Mioceno superior-Plioceno superior. Coloquios de Paleontología, 1:499-509.
Pesquero, M.D. 2003. Hipparion del Turoliense superior de Las Casiones (Fosa de Teruel). Coloquios de Paleontología, Volumen Ex:511-548.
Pesquero, M.D. and Alberdi, M.T. 2012. New evidence of conspecificity between Hipparion primigenium melendezi Alberdi, 1974 from Los Valles de Fuentidueña (Segovia) and Hipparion concudense concudense Pirlot, 1956 from Concud (Teruel) Spain. Estudios Geológicos 68:247-260. https://doi.org/10.3989/egeol.40499.152
Pesquero, M.D., Alberdi, M.T., and Montoya, P. 2007. Hipparion (Equidae, Mammalia) from Venta del Moro (Valencia Province, Spain). Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, 243:273-297. https://doi.org/10.1127/0077-7749/2007/0243-0273
Pesquero, M.D., Alcalá, L., and Fernández-Jalvo, Y. 2013. Taphonomy of the reference Miocene vertebrate mammal site of Cerro de la Garita, Spain. Lethaia 46:378-398. https://doi.org/10.1111/let.12016
Pirlot, P.L. 1956. Les Formes Européennes du genre Hipparion. Consejo Superior de Investigaciones Científicas, Barcelona.
Scott, R.S., Clavel, J., DeMiguel, D., Kaya, T., Kostopoulos, D.S., Mayda, S., and Merceron, G. 2013. Ecology of european hipparionines and the diversity of late Miocene hominids in western Eurasia, 14th RCMN Congress Neogene to Quaternary Geological Evolution of Mediterranean, Paratethys and Black Sea. Istanbul, p. 262.
Sen, S., Kondopoulou, D., and de Bonis, L. 2000. Magnetostratigraphy of Late Miocene continental deposits of the Lower Axios valley, Macedonia, Geological Society of Greece. Special Publications, 9:197-206.
Spassov, N. 2002. The Turolian megafauna of west Bulgaria and the character of the Late Miocene “Pikermian biome“. Bollettino della Società Paleontologica Italiana, 41:69-81.
Tomàs, M., Alba, D.M., Sanisidro, Ó., and Bolet, A. 2010. Los perisodáctilos del Mioceno superior de la autovía orbital de Barcelona B-40, tramo Olesa de Montserrat - Viladecavalls (Cuenca del Vallès-Penedès). Cidaris, 30:317-323.
van Dam, J. 1997. The small mammals from the Upper Miocene of the Teruel-Alfambra region (Spain): paleobiology and paleoclimatic reconstructions. Geologica Ultraiectina, 156: 1-204.
van Dam, J., Abdul Aziz, H., Álvarez Sierra, M.A., Hilgen, F.J., Van Den Hoek Ostende, L.W., Lourens, L.J., Mein, P., Van Der Meulen, A.J., and Pelaez-Campomanes, P., 2006. Long-period astronomical forcing of mammal turnover. Nature, 443:687-691. https://doi.org/10.1038/nature05163
van Dam, J.A., Alcalá, L., Zarza, A.A., Calvo, J.P., Garcés, M., and Krijgsman, W. 2001. The upper Miocene mammal record from the Teruel-Alfambra region (Spain). The MN system and continental stage/age concepts discussed. Journal of Vertebrate Paleontology, 21:367-385. https://doi.org/10.1671/0272-410.1671/0272-4634(2001)021[0367:TUMMRF]2.0.CO;2
Vlachou, T.D. 2013. Palaeontological, biostratigraphical and palaeoecological study of the Greek hipparions. PhD Thesis. Aristotle University of Thessaloniki, Greece. (in Greek with English summary).
Vlachou, T.D. and Koufos, G.D. 2009. Equidae, p. 207-281 In Koufos, G.D. and Nagel, D. (eds.), The late Miocene Mammal Faunas of the Mytilinii Basin, Samos Island, Greece: new collection. Beiträge zur Paläontologie, 31.
Woodburne, M.O. and Bernor, R.L. 1980. On superspecific groups of some Old World hipparionine horses. Journal of Paleontology, 54:1319-1348.
APPENDIX 2. Raw data of all the hipparionin sample analyzed.
Orlandi-Oliveras, Guillem (2022), “Appendices for: Feeding strategies of circum-Mediterranean hipparionins during the late Miocene: exploring dietary preferences related to size through Dental Microwear Analysis”, Mendeley Data, V1. https://data.mendeley.com/datasets/np65bh8tgw/2
APPENDIX 3. Photosimulations of the microwear surfaces.
Orlandi-Oliveras, Guillem (2022), “Appendices for: Feeding strategies of circum-Mediterranean hipparionins during the late Miocene: exploring dietary preferences related to size through Dental Microwear Analysis”, Mendeley Data, V1. https://data.mendeley.com/datasets/np65bh8tgw/2
APPENDIX 4.
Results of the Principal Component Analysis performed with the ten settings used to calculate HAsfc (4, 9, 16, 25, 36, 49, 64, 81, 100, and 121 cells). Percentage of variance explained by the principal component and the contribution of each setting to the principal components are shown.
| Component | % of variance | H4 | H9 | H16 | H25 | H36 | H49 | H64 | H81 | H100 | H121 |
| PC1 | 84.31 | 5.11 | 8.26 | 9.87 | 10.69 | 11.06 | 11.09 | 11.15 | 10.99 | 10.89 | 10.89 |
| PC2 | 7.88 | 60.94 | 15.86 | 3.57 | 0.02 | 0.62 | 2.46 | 2.68 | 4.66 | 5.18 | 4.03 |
APPENDIX 5. Metapodial measures and body mass estimations.
Orlandi-Oliveras, Guillem (2022), “Appendices for: Feeding strategies of circum-Mediterranean hipparionins during the late Miocene: exploring dietary preferences related to size through Dental Microwear Analysis”, Mendeley Data, V1. https://data.mendeley.com/datasets/np65bh8tgw/2
APPENDIX 6. Linear regressions on DMT parameters versus body mass.
APPENDIX 6.1. Linear regressions of microwear textural parameters and the calculated principal component against body mass for all hipparionins. Correlation statistics included in each graph. A: Regression of the Asfc values. B: epLsar. C: HAsfc. D: Tfv. E: PC1.
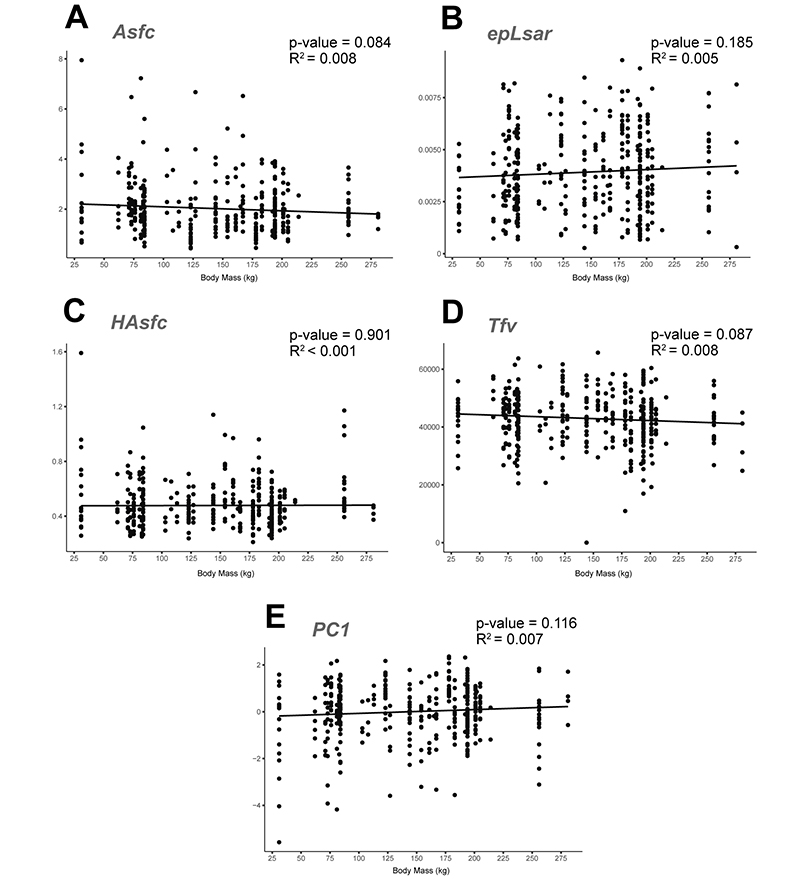
APPENDIX 6.2. Linear regressions of microwear textural parameters and the calculated principal component against body mass for western Mediterranean hipparionins. Correlation statistics included in each graph. A: Regression of the Asfc values. B: epLsar. C: HAsfc. D: Tfv. E: PC1.
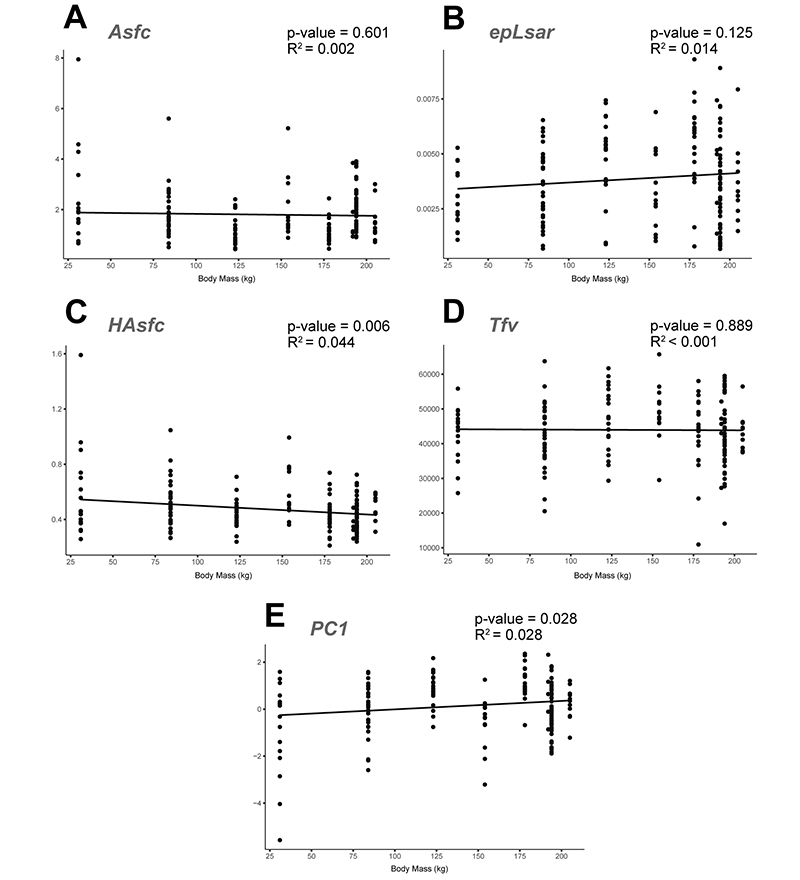
APPENDIX 6.3. Linear regressions of microwear textural parameters and the calculated principal component against body mass for hipparionins from the Balkans. Correlation statistics included in each graph. A: Regression of the Asfc values. B: epLsar. C: HAsfc. D: Tfv. E: PC1.
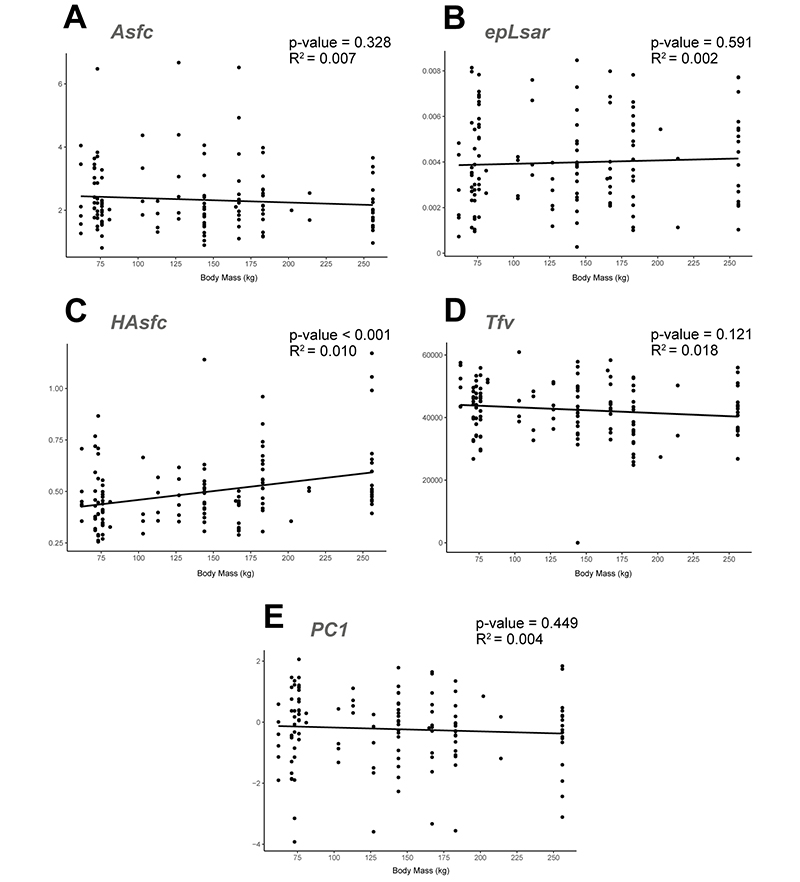
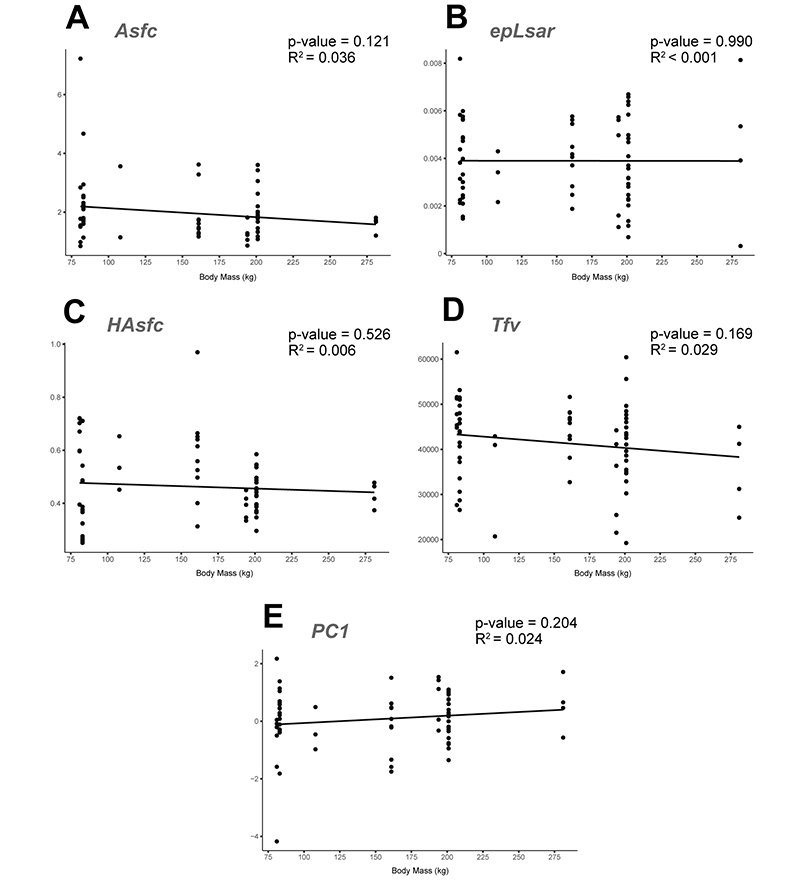
APPENDIX 6.4. Linear regressions of microwear textural parameters and the calculated principal component against body mass for western Anatolian hipparionins. Correlation statistics included in each graph. A: Regression of the Asfc values. B: epLsar. C: HAsfc. D: Tfv. E: PC1.
APPENDIX 7. Post-hoc pairwise comparisons with Turolian hipparionin data.
APPENDIX 7.1. Post-hoc tests on ranked complexity (Asfc). P-values from HSD test are depicted above the grey diagonal and those from LSD are under it. Significant p-values represented in bold.
| BIOPROVINCE | Western Mediterranean | Balkans | Western Anatolia |
| Western Mediterranean | < 0.001 | 0.001 | |
| Balkans | < 0.001 | 0.011 | |
| Western Anatolia | < 0.001 | 0.004 |
APPENDIX 7.2. Post-hoc tests on ranked heterogeneity of complexity (HAsfc). P-values from HSD test are depicted above the grey diagonal and those from LSD are under it. Significant p-values represented in bold.
| BIOPROVINCE | Western Mediterranean | Balkans | Western Anatolia |
| Western Mediterranean | 0.049 | 0.935 | |
| Balkans | 0.019 | 0.189 | |
| Western Anatolia | 0.727 | 0.081 |
APPENDIX 7.3. Summary of post-hoc tests on ranked parameters showing significant differences between the Turolian hipparionin groups. Significant differences between groups obtained from HSD test are depicted above the grey diagonal and those from LSD are under it.
| Bioprovince | West Mediterranean | Balkans | Western Anatolia | ||||||||||
| Turolian hipparionin groups | periafricanum | gromovae | cf. matthewi | concudense | primigenium | macedonicum | dietrichi | proboscideum | primigenium | macedonicum | dietrichi | proboscideum | |
| Western Mediterranean | periafricanum | epLsar | Asfc, epLsar, PC1 | ||||||||||
| gromovae | Asfc, epLsar, PC1 | Asfc | |||||||||||
| cf. matthewi | Asfc, epLsar, PC1 | Asfc, epLsar, PC1 | Asfc | Asfc | Asfc, PC1 | Asfc, PC1 | Asfc, PC1 | Asfc | |||||
| concudense | Asfc, epLsar, PC1 | Asfc, epLsar, PC1 | Asfc, PC1 | Asfc, PC1 | Asfc, PC1 | Asfc, HAsfc, PC1 | Asfc, PC1 | PC1 | Asfc, PC1 | ||||
| primigenium | HAsfc | HAsfc | HAsfc | ||||||||||
| Balkans | macedonicum | Asfc, epLsar | Asfc, | Asfc, PC1 | Asfc, epLsar, PC1 | Asfc, | HAsfc | ||||||
| dietrichi | Asfc, | Asfc, epLsar, PC1 | Asfc, epLsar, PC1 | HAsfc, | |||||||||
| proboscideum | epLsar, PC1 | Asfc, HAsfc, PC1 | Asfc, HAsfc, PC1 | Asfc, epLsar, HAsfc, PC1 | Asfc, HAsfc, PC1 | HAsfc, PC1 | PC1 | HAsfc | |||||
| primigenium | epLsar | HAsfc | Asfc, HAsfc, PC1 | Asfc, epLsar, HAsfc, PC1 | HAsfc | HAsfc | HAsfc | HAsfc | |||||
| Western Anatolia | macedonicum | Asfc, PC1 | Asfc, epLsar, PC1 | HAsfc | HAsfc | ||||||||
| dietrichi | Asfc, HAsfc, PC1 | Asfc, epLsar, HAsfc, PC1 | HAsfc | Asfc, HAsfc | Asfc | HAsfc | |||||||
| proboscideum | epLsar | Asfc, PC1 | Asfc, epLsar, PC1 | Asfc | Asfc, HAsfc, PC1 | HAsfc | HAsfc | ||||||
APPENDIX 7.4. Summary of post-hoc tests on parameters showing significant differences between localities. Here represented the results from pairwise comparisons between localities with hipparionins of the macedonicum group. Significant differences obtained from the HSD test are above the grey diagonal and those from LSD are under it.
| macedonicum group | Balkans | Western Anatolia | ||||||
| Locality | NIK | RZO | PER | DTK | GLP | MYSE | MTLA/B | |
| Balkans | Nikiti-2 (NIK) | epLsar | ||||||
| Ravin des Zouaves-5 (RZO) | epLsar | |||||||
| Perivolaki (PER) | epLsar, PC1 | |||||||
| Dytiko (DTK) | epLsar, PC1 | Tfv | ||||||
| Western Anatolia | Gülpınar (GLP) | HAsfc, PC1 | HAsfc | HAsfc | HAsfc | |||
| Şerefköy-2 (MYSE) | epLsar | Tfv | HAsfc | |||||
| Samos (MTLA/B) | Tfv | HAsfc | ||||||
APPENDIX 7.5. Summary of post-hoc tests on parameters showing significant differences between localities. Here represented the results from pairwise comparisons between localities with hipparionins of the dietrichi group. Significant differences obtained from the HSD test are above the grey diagonal and those from LSD are under it.
| dietrichi group | Balkans | Western Anatolia | ||||
| Locality | NIK | PER | DTK | GLP | MTLA/B | |
| Balkans | Nikiti-2 (NIK) | |||||
| Perivolaki (PER) | epLsar, PC1 | |||||
| Dytiko (DTK) | ||||||
| Western Anatolia | Gülpınar (GLP) | |||||
| Samos (MTLA/B) | epLsar | |||||
APPENDIX 7.6. Summary of post-hoc tests on parameters showing significant differences between localities. Here represented the results from pairwise comparisons between localities with hipparionins of the proboscideum group. Significant differences obtained from the HSD test are above the grey diagonal and those from LSD are under it.
| proboscideum group | Balkans | Western Anatolia | ||||
| Locality | RZO | PER | HD | MYSE | MTLA/B | |
| Balkans | Ravin des Zouaves-5 (RZO) | |||||
| Perivolaki (PER) | Tfv | |||||
| Hadjidimovo (HD) | ||||||
| Western Anatolia | Şerefköy-2 (MYSE) | HAsfc | ||||
| Samos (MTLA/B) | Tfv, PC1 | HAsfc, PC1 | Tfv | |||
APPENDIX 7.7. Summary of post-hoc tests on parameters showing significant differences between localities. Here represented the results from pairwise comparisons between localities with hipparionins of the primigenium group. Significant differences obtained from the HSD test are above the grey diagonal and those from LSD are under it.
| primigenium group | Western Mediterranean | Balkans | |
| Locality | ARQ | HD | |
| Western Mediterranean | El Arquillo (ARQ) | HSD test → | |
| Balkans | Hadjidimovo (HD) | HAsfc | ← LSD test |
APPENDIX 7.8. Results of the post-hoc tests from pairwise comparisons between hipparionin groups from Nikiti-2 locality.
| Nikiti-2 (NIK) | macedonicum | dietrichi |
| macedonicum | HSD test → | Asfc: - |
| epLSar: 1.000 | ||
| HAsfc: 1.000 | ||
| Tfv: 1.000 | ||
| PC1: 1.000 | ||
| dietrichi | Asfc: - | ← LSD test |
| epLSar: 0.968 | ||
| HAsfc: 0.480 | ||
| Tfv: 0.667 | ||
| PC1: 0.811 |
APPENDIX 7.9. Results of the post-hoc tests from pairwise comparisons between hipparionin groups from Perivolaki locality.
| Perivolaki (PER) | macedonicum | dietrichi | proboscideum |
| macedonicum | HSD test → | Asfc: - | Asfc: - |
| epLSar: 1.000 | epLSar: 1.000 | ||
| HAsfc: 1.000 | HAsfc: 1.000 | ||
| Tfv: 1.000 | Tfv: 1.000 | ||
| PC1: 1.000 | PC1: 1.000 | ||
| dietrichi | Asfc: - |
|
Asfc: - |
| epLSar: 0.813 | epLSar: 1.000 | ||
| HAsfc: 0.569 | HAsfc: 1.000 | ||
| Tfv: 0.997 | Tfv: 1.000 | ||
| PC1: 0.389 | PC1: 1.000 | ||
| proboscideum | Asfc: - | Asfc: - | ← LSD test |
| epLSar: 0.744 | epLSar: 0.669 | ||
| HAsfc: 0.779 | HAsfc: 0.997 | ||
| Tfv: 0.184 | Tfv: 0.198 | ||
| PC1: 0.203 | PC1: 0.408 |
APPENDIX 7.10. Results of the post-hoc tests from pairwise comparisons between hipparionin groups from Hadjidimovo locality.
| Hadjidimovo (HD) | proboscideum | primigenium |
| proboscideum | HSD test → | Asfc: - |
| epLSar: 1.000 | ||
| HAsfc: 1.000 | ||
| Tfv: 1.000 | ||
| PC1: 1.000 | ||
| primigenium | Asfc: - | ← LSD test |
| epLSar: 0.970 | ||
| HAsfc: 0.990 | ||
| Tfv: 0.257 | ||
| PC1: 0.187 |
APPENDIX 7.11. Results of the post-hoc tests from pairwise comparisons between hipparionin groups from Dytiko locality. Significant p-values represented in bold.
| Dytiko (DTK) | macedonicum | dietrichi |
| macedonicum | HSD test → | Asfc: - |
| epLSar: 0.998 | ||
| HAsfc: 1.000 | ||
| Tfv: 0.913 | ||
| PC1: 1.000 | ||
| dietrichi | Asfc: - | ← LSD test |
| epLSar: 0.144 | ||
| HAsfc: 0.806 | ||
| Tfv: 0.040 | ||
| PC1: 0.323 |
APPENDIX 7.12. Results of the post-hoc tests from pairwise comparisons between hipparionin groups from Gülpınar locality.
| Gülpınar (GLP) | macedonicum | dietrichi |
| macedonicum | HSD test → | Asfc: - |
| epLSar: 1.000 | ||
| HAsfc: 1.000 | ||
| Tfv: 1.000 | ||
| PC1: 1.000 | ||
| dietrichi | Asfc: - | ← LSD test |
| epLSar: 0.991 | ||
| HAsfc: 0.448 | ||
| Tfv: 0.444 | ||
| PC1: 0.516 |
APPENDIX 7.13. Results of the post-hoc tests from pairwise comparisons between hipparionin groups from Samos new collections.
| Samos (MTLA/B) | macedonicum | dietrichi | proboscideum |
| macedonicum | HSD → | Asfc: - | Asfc: - |
| epLSar: 1.000 | epLSar: 1.000 | ||
| HAsfc: 1.000 | HAsfc: 0.971 | ||
| Tfv: 1.000 | Tfv: 1.000 | ||
| PC1: 1.000 | PC1: 0.995 | ||
| dietrichi | Asfc: - | Asfc: - | |
| epLSar: 0.663 | epLSar: 1.000 | ||
| HAsfc: 0.353 | HAsfc: 1.000 | ||
| Tfv: 0.850 | Tfv: 1.000 | ||
| PC1: 0.691 | PC1: 1.000 | ||
| proboscideum | Asfc: - | Asfc: - | ← LSD |
| epLSar: 0.475 | epLSar: 0.213 | ||
| HAsfc: 0.066 | HAsfc: 0.479 | ||
| Tfv: 0.902 | Tfv: 0.723 | ||
| PC1: 0.111 | PC1: 0.267 |
APPENDIX 7.14. Results of the post-hoc tests from pairwise comparisons between hipparionin groups from Şerefköy-2 locality.
| Şerefköy-2 (MYSE) | macedonicum | proboscideum |
| macedonicum | HSD test → | Asfc: - |
| epLSar: 1.000 | ||
| HAsfc: 0.997 | ||
| Tfv: 1.000 | ||
| PC1: 1.000 | ||
| proboscideum | Asfc: - | ← LSD test |
| epLSar: 0.972 | ||
| HAsfc: 0.124 | ||
| Tfv: 0.881 | ||
| PC1: 0.659 |
APPENDIX 7.15. Results of the post-hoc tests from pairwise comparisons between hipparionin groups from El Arquillo locality. Significant p-values represented in bold.
| El Arquillo (ARQ) | periafricanum | gromovae | primigenium |
| periafricanum | HSD → | Asfc: - | Asfc: - |
| epLSar: 1.000 | epLSar: 0.995 | ||
| HAsfc: 1.000 | HAsfc: 0.840 | ||
| Tfv: 1.000 | Tfv: 1.000 | ||
| PC1: 1.000 | PC1: 1.000 | ||
| gromovae | Asfc: - | Asfc: - | |
| epLSar: 0.175 | epLSar: 1.000 | ||
| HAsfc: 0.645 | HAsfc: 0.909 | ||
| Tfv: 0.779 | Tfv: 0.985 | ||
| PC1: 0.743 | PC1: 1.000 | ||
| primigenium | Asfc: - | Asfc: - | ← LSD |
| epLSar: 0.112 | epLSar: 0.397 | ||
| HAsfc: 0.027 | HAsfc: 0.039 | ||
| Tfv: 0.970 | Tfv: 0.830 | ||
| PC1: 0.604 | PC1: 0.450 |
