

APPENDIX 1.
Phylogenetic relationships between †Otodontidae and Lamnidae proposed by Greenfield (2022a, 2022b).
Greenfield (2022a, 2022b) made new propositions regarding the close phylogenetic affinity between †Otodontidae and Lamnidae and presented a body restoration of another otodontid taxon. Although they are not central to the scope of this present paper, Greenfield’s (2022a, 2022b) propositions are relevant to the context of either Cooper et al.’s (2022) phylogenetic assertion in relation to †O. megalodon or considering the body form of †O. megalodon and otodontids in general. Therefore, we briefly comment on Greenfield’s (2022a, 2022b) propositions here.
Phylogenetic Position of †O. megalodon
Cooper et al. (2022) noted that “there are uncertainties regarding the interrelationships between extinct and extant Lamniformes, and therefore, † O. megalodon could be as closely related to C. carcharias as to any other lamniform” (p. 9). Subsequently, Greenfield (2022a, 2022b) contended that †Otodontidae is a sister to the family Lamnidae and even erected a new superfamily Lamnoidea that represents a clade consisting of †Otodontidae and Lamnidae. Greenfield (2022a, 2022b) argued that the following two characters phylogenetically unite †Otodontidae and Lamnidae: 1) robust, calcified rostral cartilages with circular transverse cross-section and without fenestrae and appendices; and 2) regional endothermy. It is important to point out that the taxonomic identity of the isolated fossil rostral cartilages used for Greenfield’s (2022a, 2022b) basis remains inferential and requires testing by the discovery of rostral specimens associated with taxonomically diagnostic teeth from the same individual shark. We also note that rostral hypercalcification is not confined to Lamnidae (more specifically Lamna: Figure 3) within Lamniformes but also occurs in the extant bigeye thresher (Alopias superciliosus: Figure A1) and possibly at least in one member of the Cretaceous taxon, †Anacoracidae (Shimada, 2013). In addition, the assertion that regional endothermy can be used as a synapomorphy to unite †Otodontidae and Lamnidae within Lamniformes is now questionable based on the recent papers suggesting that regional endothermy may be present broadly within Lamniformes (Dolton et al., 2023a, 2023b, and references therein; but see also Griffiths et al., 2023). Greenfield (2022b) noted whether †Otodontidae lies outside of the ‘Cetorhinidae+Lamnidae’ clade has never been tested. However, it should be pointed out that there has been no test conducted on or supporting the ‘†Otodontidae+Lamnidae’ clade either, unlike the existing strong support of the sister relationship between Cetorhinidae and Lamnidae both morphologically and molecularly (Stone and Shimada, 2019; Vella and Vella, 2020; Kousteni et al., 2021; Silva et al., 2023). Even if Cetorhinidae and Lamnidae are not sisters hypothetically (vs. Figure 1A), it will not automatically place †Otodontidae as a sister to Lamnidae simply because there is so far not even one decisive synapomorphy that supports Greenfield’s (2022a, 2022b) ‘Lamnoidea hypothesis.’ We stress that the practice of erecting higher taxonomic categories should be conducted carefully, especially for a phylogenetically complex group like Lamniformes (Stone and Shimada, 2019; Shimada, 2022).
Another Problematic Otodontid Body Restoration
Greenfield (2022b) presented a body restoration of another otodontid, † Cretalamna sp., based on previously published photographs of multiple partial or nearly complete skeletal specimens from the Upper Cretaceous (Cenomanian Stage) of Lebanon (Pfeil, 2021). Greenfield (2022b) used his restoration to further support his ‘Lamnoidea hypothesis’ on the basis of its putative body form that is vaguely reminiscent of extant lamnids, particularly Lamna. Such a restoration of another otodontid may appear significantly relevant to the context of restoring the body form of †O. megalodon. However, Greenfield’s (2022b) †Cretalamna body restoration must be viewed with skepticism. This is because, while the degree of taphonomic distortion or flattening of the body through the fossilization of the specimens was not assessed, the authenticity of the specimens or anatomical parts of the specimens can only be speculated from the photographs. More critically, the † Cretalamna skeletons (Pfeil, 2021) are ‘unnumbered’ specimens with uncertain catalog status. Effectively, this condition does not allow the reproducibility of Greenfield’s (2022b) propositions, and thus, they cannot be viewed as scientifically valid.
REFERENCES
Cooper, J.A., Hutchinson, J.R., Bernvi, D.C., Cliff, G., Wilson, R.P., Dicken, M.L., Menzel, J., Wroe, S, Pirlo, J., and Pimiento, C. 2022. The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling. Science Advances, 8:eabm9424.
https://doi.org/10.1126/sciadv.abm9424
Dolton, H.R., Jackson, A.L., Deaville, R., Hall, J., Hall, G., McManus G., Perkins, M.R., Rolfe, R.A., Snelling, E.P., Houghton, J.D.R., Sims, D.W., and Payne, N.L. 2023a. Regionally endothermic traits in the planktivorous basking sharks Cetorhinus maximus. Endangered Species Research, 51:227-232.
https://doi.org/10.3354/esr01257
Dolton, H.R., Snelling, E.P., Deaville, R., Jackson, A.L., Perkins, M.W., Bortoluzzi, J.R., Purves, K., Curnick, D.J., Pimiento, C., and Payne, N.L. 2023b. Centralized red muscle in Odontaspis ferox and the prevalence of regional endothermy in sharks. Biology Letters, 19:20230331.
https://doi.org/10.1098/rsbl.2023.0331
Greenfield, T. 2022a. List of skeletal material from megatooth sharks (Lamniformes: Otodontidae). Paleoichthys, 4:1-9.
Greenfield, T. 2022b. Additions to “List of skeletal material from megatooth sharks”, with a response to Shimada (2022). Paleoichthys, 6:6-11.
Griffths, M.L., Eagle, R.A., Kim, S.L., Flores, R.J., Becker, M.A., Maisch IV, H.M., Trayler, R.B., Chan, R.L., McCormack J., Akhtar, A.A., Tripati, A.K., and Shimada, K. 2023. Endothermic physiology of extinct megatooth sharks. Proceedings of the National Academy of Sciences, 120:e221815312.
https://doi.org/10.1073/pnas.2218153120
Kousteni, V., Mazzoleni, S., Vasileidadou, K., and Rovatsos, M. 2021. Complete mitochondrial DNA genome of nine species of sharks and rays and their phylogenetic placement among modern elasmobranchs. Genes, 12:324.
https://doi.org/10.3390/genes12030324
Pfeil, F.H. 2021. The new family Mesiteiidae (Chondrichthyes, Orectolobiformes) based on Mesiteida emiliae Kramberger, 1884. A contribution to the Upper Cretaceous (early Cenomanian) shark fauna from Lebanon. In Pradel, A., Denton, J.S.S., and Janvier, P. (eds.) Ancient Fishes and Their Living Relatives: A Tribute to John G. Maisey. Verlag Dr. Friedrich Pfeil, Munich, DE.
Shimada, K. 2013. Chondrichthyan origin for the fossil record of the tselfatiiform osteichthyan fish, Thryptodus zitteli Loomis, from the Upper Cretaceous Mooreville Chalk of Alabama. Bulletin of the Alabama Museum of Natural History, 31:72-77.
Shimada, K. 2022. Phylogenetic affinity of the extinct shark family Otodontidae within Lamniformes remains uncertain–Comments on “List of skeletal material from megatooth sharks (Lamniformes, Otodontidae)” by Greenfield. Paleoichthys, 6:1-5.
Silva, J.P.C.B, Shimada, K., and Datovo, A. 2023. The importance of the appendicular skeleton for the phylogenetic reconstruction of lamniform sharks (Chondrichthyes: Elasmobranchii). Journal of Morphology, 284:e21585.
https://doi.org/10.1002/jmor.21585
Stone, N. R. and Shimada, K. 2019. Skeletal anatomy of the bigeye sandtiger shark, Odontaspis noronhai (Lamniformes: Odontaspididae), and its implications for lamniform phylogeny, taxonomy, and conservation biology. Copeia, 107: 632-652.
https://doi.org/10.1643/CG-18-160
Vella, N. and Vella, A. 2020. The complete mitogenome of the critically endangered smalltooth sand tiger shark, Odontaspis ferox (Lamniformes: Odontaspididae). Mitochondrial DNA B Resources, 5:3301-3304.
https://doi.org/10.1080/23802359.2020.1814886
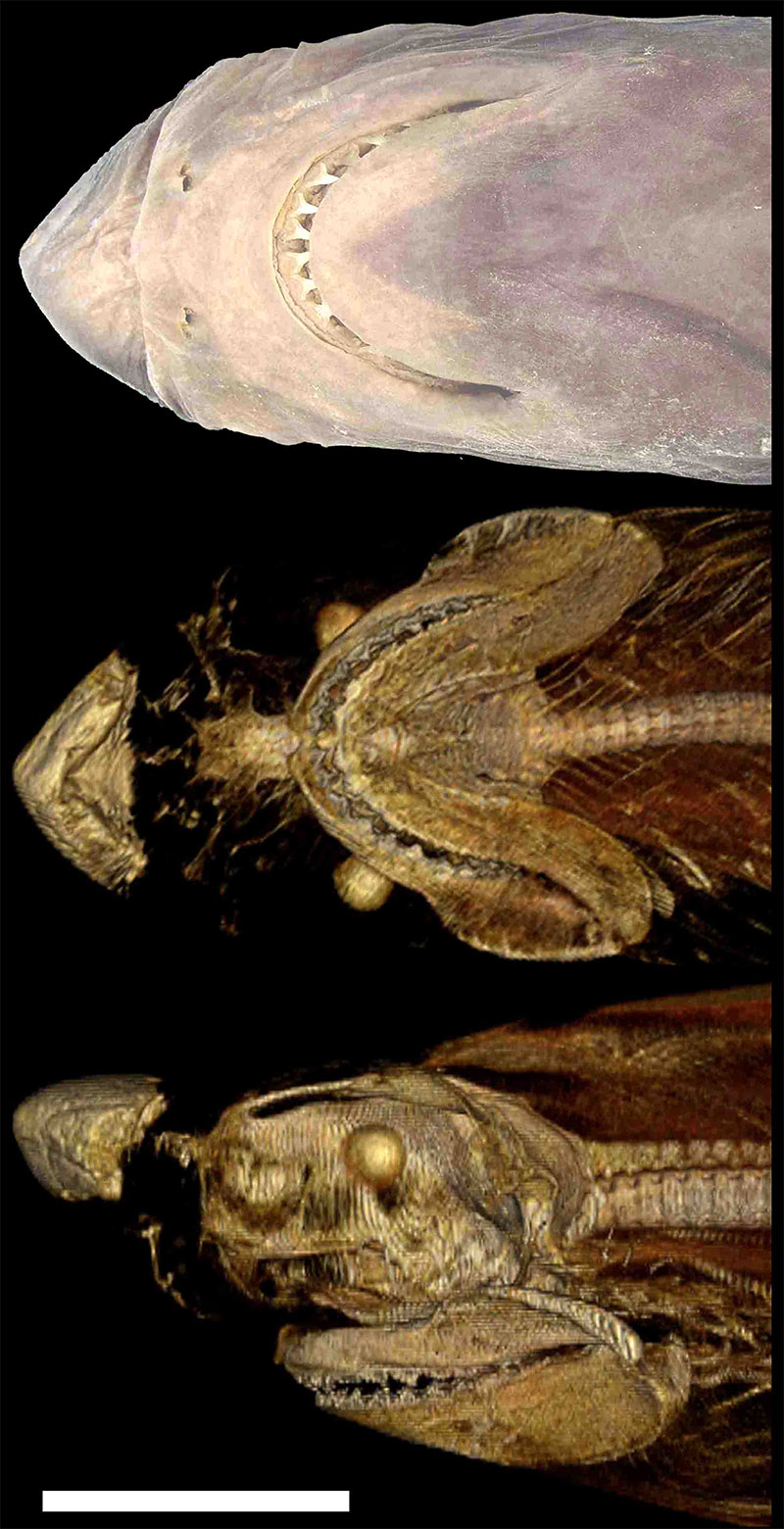
FIGURE A1. Photographic (*) and CT (**) images of cranial region of 187-cm-TL male extant bigeye thresher (Alopias superciliosus: UF 160188) caught off Florida, USA, demonstrating hypercalcified rostral cartilage in the species. Top, ventral view*; middle, ventral view**; bottom, left lateral view **. Note that the same hypercalcification is also present in another specimen of A. superciliosus (UF 178509: 201-cm-TL male caught off Florida). Scale bar = 10 cm.
APPENDIX 2.
Measurements of the Carcharodon carcharias specimen LACM 43805-1.
| Vertebra | Ø #1 (mm) | Ø #2 (mm) | Ø #3 (mm) | Ø mean (mm) |
|---|---|---|---|---|
| #1 | 12.73 | 12.45 | 12.67 | 12.62 |
| #2 | 13.41 | 13.08 | 13.15 | 13.21 |
| #3 | 13.81 | 13.44 | 13.51 | 13.59 |
| #4 | 13.77 | 13.45 | 13.56 | 13.59 |
| #5 | 14.54 | 13.88 | 14.39 | 14.27 |
| #6 | 14.73 | 13.58 | 14.52 | 14.27 |
| #7 | 14.65 | 14.05 | 14.06 | 14.25 |
| #8 | 14.90 | 14.15 | 14.02 | 14.36 |
| #9 | 14.56 | 14.67 | 14.48 | 14.57 |
| #10 | 15.17 | 15.15 | 15.22 | 15.18 |
| #11 | 15.41 | 15.24 | 15.53 | 15.39 |
| #12 | 16.23 | 15.34 | 15.75 | 15.77 |
| #13 | 15.65 | 15.04 | 15.86 | 15.52 |
| #14 | 16.42 | 16.21 | 16.22 | 16.28 |
| #15 | 16.52 | 15.92 | 16.1 | 16.18 |
| #16 | 15.99 | 16.1 | 16.2 | 16.10 |
| #17 | 16.09 | 16.19 | 15.92 | 16.07 |
| #18 | 16.26 | 16.79 | 16.04 | 16.36 |
| #19 | 16.38 | 16.66 | 16.20 | 16.41 |
| #20 | 16.27 | 16.42 | 16.13 | 16.27 |
| #21 | 16.23 | 16.36 | 16.12 | 16.24 |
| #22 | 16.38 | 16.38 | 16.30 | 16.35 |
| #23 | 16.48 | 16.43 | 16.71 | 16.54 |
| #24 | 16.65 | 16.38 | 16.5 | 16.51 |
| #25 | 16.91 | 16.19 | 16.99 | 16.70 |
| #26 | 16.77 | 16.38 | 16.17 | 16.44 |
| #27 | 16.71 | 16.02 | 16.4 | 16.38 |
| #28 | 16.83 | 16.38 | 16.1 | 16.44 |
| #29 | 17.24 | 17.09 | 16.54 | 16.96 |
| #30 | 17.17 | 16.9 | 17.02 | 17.03 |
| #31 | 17.08 | 17.18 | 17.27 | 17.18 |
| #32 | 17.24 | 17.63 | 17.43 | 17.43 |
| #33 | 17.36 | 16.78 | 17.52 | 17.22 |
| #34 | 17.9 | 17.22 | 17.73 | 17.62 |
| #35 | 17.92 | 17.82 | 17.71 | 17.82 |
| #36 | 18.16 | 17.89 | 17.82 | 17.95 |
| #37 | 17.95 | 17.37 | 17.73 | 17.68 |
| #38 | 18.33 | 17.75 | 17.83 | 17.97 |
| #39 | 18.44 | 18.17 | 17.92 | 18.18 |
| #40 | 18.38 | 17.91 | 17.85 | 18.05 |
| #41 | 18.79 | 18.08 | 18.13 | 18.33 |
| #42 | 18.29 | 17.97 | 18.22 | 18.16 |
| #43 | 18.07 | 17.79 | 18.09 | 17.98 |
| #44 | 18.38 | 18.03 | 18.17 | 18.19 |
| #45 | 18.73 | 18.10 | 18.2 | 18.34 |
| #46 | 18.46 | 18.25 | 18.44 | 18.38 |
| #47 | 18.86 | 18.13 | 18.29 | 18.43 |
| #48 | 18.97 | 18.69 | 19.09 | 18.92 |
| #49 | 19.31 | 18.93 | 18.52 | 18.92 |
| #50 | 19.19 | 18.97 | 18.71 | 18.96 |
| #51 | 18.73 | 18.41 | 18.71 | 18.62 |
| #52 | 19.03 | 18.51 | 18.32 | 18.62 |
| #53 | 18.98 | 18.63 | 18.57 | 18.73 |
| #54 | 19.46 | 18.71 | 19.05 | 19.07 |
| #55 | 19.53 | 19.08 | 19.33 | 19.31 |
| #56 | 19.6 | 19.43 | 19.23 | 19.42 |
| #57 | 19.53 | 18.43 | 19.38 | 19.11 |
| #58 | 19.31 | 19.14 | 19.06 | 19.17 |
| #59 | 19.24 | 19.65 | 19.75 | 19.55 |
| #60 | 19.19 | 19.61 | 19.56 | 19.45 |
| #61 | 19.47 | 19.94 | 19.85 | 19.75 |
| #62 | 18.79 | 18.91 | 20.05 | 19.25 |
| #63 | 19.2 | 19.54 | 19.61 | 19.45 |
| #64 | 18.57 | 19.18 | 19.31 | 19.02 |
| #65 | 17.9 | 18.45 | 18.73 | 18.36 |
| #66 | 17.94 | 18.14 | 18.3 | 18.13 |
| #67 | 18.87 | 17.83 | 18.28 | 18.33 |
| #68 | 18.48 | 18.55 | 18.18 | 18.40 |
| #69 | 18.93 | 18.55 | 18.44 | 18.64 |
| #70 | 18.59 | 18.1 | 18.04 | 18.24 |
| #71 | 18.51 | 18.44 | 18.41 | 18.45 |
| #72 | 18.93 | 18.67 | 18.28 | 18.63 |
| #73 | 18.96 | 18.65 | 18.53 | 18.71 |
| #74 | 18.36 | 18.26 | 18.44 | 18.35 |
| #75 | 19.28 | 17.65 | 18.12 | 18.35 |
| #76 | 17.77 | 18.79 | 18.82 | 18.46 |
| #77 | 17.18 | 17.46 | 18.36 | 17.67 |
| #78 | 18.21 | 18.12 | 18.22 | 18.18 |
| #79 | 17.09 | 17.29 | 17.67 | 17.35 |
| #80 | 17.15 | 17.18 | 17.38 | 17.24 |
| #81 | 16.39 | 16.88 | 16.84 | 16.70 |
| #82 | 16.08 | 16.66 | 16.58 | 16.44 |
| #83 | 15.99 | 16.47 | 16.4 | 16.29 |
| #84 | 16.1 | 16.38 | 16.44 | 16.31 |
| #85 | 15.67 | 16.23 | 16.02 | 15.97 |
| #86 | 15.45 | 15.66 | 16.21 | 15.77 |
| #87 | 15.2 | 15.48 | 15.81 | 15.50 |
| #88 | 15.16 | 15.28 | 15.2 | 15.21 |
| #89 | 14.87 | 15.04 | 15.45 | 5.12 |
| #90 | 15.38 | 15.37 | 15.53 | 15.43 |
| #91 | 14.39 | 15.32 | 15.61 | 15.11 |
| #92 | 14.62 | 15.54 | 15.05 | 15.07 |
| #93 | 14.28 | 14.87 | 14.69 | 14.61 |
| #94 | 13.71 | 14.15 | 14.3 | 14.05 |
| #95 | 13.63 | 13.77 | 13.56 | 13.65 |
| #96 | 13.79 | 13.77 | 14.06 | 13.87 |
| #97 | 13.98 | 13.63 | 13.77 | 13.79 |
| #98 | 13.79 | 13.6 | 13.66 | 13.68 |
| #99 | 13.45 | 13.55 | 13.46 | 13.49 |
| #100 | 13.36 | 13.51 | 13.53 | 13.47 |
| #101 | 13.1 | 13.11 | 13.02 | 13.08 |
| #102 | 12.81 | 12.33 | 12.97 | 12.70 |
| #103 | 12.49 | 12.12 | 12.81 | 12.47 |
| #104 | 12.52 | 12.02 | 12.31 | 12.28 |
| #105 | 12.23 | 11.82 | 12.05 | 12.03 |
| #106 | 12.36 | 11.82 | 12.12 | 12.10 |
| #107 | 12.24 | 11.82 | 11.71 | 11.92 |
| #108 | 11.97 | 12.12 | 12.14 | 12.08 |
| #109 | 12.21 | 11.44 | 11.75 | 11.80 |
| #110 | 11.77 | 11.18 | 11.71 | 11.55 |
| #111 | 11.78 | 11.18 | 11.41 | 11.46 |
| #112 | 11.47 | 10.94 | 11.38 | 11.26 |
| #113 | 10.93 | 10.62 | 10.88 | 10.81 |
| #114 | 10.33 | 10.41 | 10.45 | 10.40 |
| #115 | 10.36 | 9.98 | 10.45 | 10.26 |
| #116 | 10.11 | 10.05 | 10.16 | 10.11 |
| #117 | 9.79 | 10.09 | 9.87 | 9.92 |
| #118 | 9.55 | 9.91 | 9.32 | 9.59 |
| #119 | 9.46 | 9.74 | 8.66 | 9.29 |
| #120 | 8.99 | 9.26 | 8.99 | 9.08 |
| #121 | 8.84 | 9.66 | 8.83 | 9.11 |
| #122 | 8.5 | 8.78 | 8.05 | 8.44 |
| #123 | 8.93 | 9.06 | 9.08 | 9.02 |
| #124 | 8.51 | 8.91 | 8.80 | 8.74 |
| #125 | 8.49 | 9.39 | 9.61 | 9.16 |
| #126 | 7.88 | 8.61 | 8.54 | 8.34 |
| #127 | 7.93 | 8.83 | 8.43 | 8.40 |
| #128 | 7.34 | 7.96 | 8.18 | 7.83 |
| #129 | 7.73 | 7.97 | 7.78 | 7.83 |
| #130 | 7.2 | 7.85 | 7.16 | 7.40 |
| #131 | 7.08 | 7.19 | 6.92 | 7.06 |
| #132 | 7.2 | 7.73 | 6.87 | 7.27 |
| #133 | 6.8 | 7.11 | 6.82 | 6.91 |
| #134 | 6.41 | 7.03 | 6.58 | 6.67 |
| #135 | 6.4 | 6.37 | 6.07 | 6.28 |
| #136 | 6.49 | 7.11 | 6.44 | 6.68 |
| #137 | 6.58 | 7.02 | 6.14 | 6.58 |
| #138 | 6.4 | 6.35 | 5.81 | 6.19 |
| #139 | 6.08 | 6.07 | 5.82 | 5.99 |
| #140 | 5.78 | 5.96 | 5.82 | 5.85 |
| #141 | 5.62 | 5.87 | 5.32 | 5.60 |
| #142 | 5.76 | 5.63 | 5.26 | 5.55 |
| #143 | 5.78 | 5.76 | 5.38 | 5.64 |
| #144 | 5 | 5.32 | 4.97 | 5.10 |
| #145 | 5.18 | 5.34 | 4.68 | 5.07 |
| #146 | 4.86 | 5.05 | 4.69 | 4.87 |
| #147 | 4.72 | 4.87 | 4.58 | 4.72 |
| #148 | 4.42 | 5.03 | 4.52 | 4.66 |
| #149 | 4.23 | 5.16 | 4.34 | 4.58 |
| #150 | 4.23 | 4.65 | 4.12 | 4.33 |
| #151 | 4.37 | 4.12 | 3.85 | 4.11 |
| #152 | 3.64 | 3.84 | 3.79 | 3.76 |
| #153 | 3.84 | 4.35 | 4.27 | 4.15 |
| #154 | 3.72 | 4.34 | 4 | 4.02 |
| #155 | 3.2 | 3.97 | 3.54 | 3.57 |
| #156 | 2.83 | 3.41 | 3.26 | 3.17 |
| #157 | 2.86 | 3.49 | 3.22 | 3.19 |
| #158 | 2.44 | 3.26 | 3.07 | 2.92 |
| #159 | 2.56 | 2.94 | 2.99 | 2.83 |
| #160 | 2.21 | 2.82 | 2.99 | 2.67 |
| #161 | 2.21 | 2.21 | 2.61 | 2.34 |
| #162 | 2.56 | 2.81 | 2.84 | 2.74 |
| #163 | 2 | 2.33 | 2.17 | 2.17 |
| #164 | 1.74 | 2.05 | 1.86 | 1.88 |
| #165 | 1.89 | 1.92 | 1.95 | 1.92 |
| #166 | ? | ? | ? | ? |
| #167 | ? | ? | ? | ? |
| #168 | ? | ? | ? | ? |
| #169 | ? | ? | ? | ? |
| #170 | ? | ? | ? | ? |
| #171 | ? | ? | ? | ? |
